

![]() A primeira coisa que pensamos quando lemos o termo “autotrófico” é em plantas. No máximo em microalgas. Alguns pensariam que também existem bactérias que fazem fotossíntese e se encaixariam nesta classificação. Mas vamos tentar entender a origem deste termo.
A primeira coisa que pensamos quando lemos o termo “autotrófico” é em plantas. No máximo em microalgas. Alguns pensariam que também existem bactérias que fazem fotossíntese e se encaixariam nesta classificação. Mas vamos tentar entender a origem deste termo.
Autotróficos seriam os organismos capazes de se “auto nutrir”, produzir moléculas orgânicas a partir de moléculas inorgânicas e da energia luminosa. Certo? Bem, não tão certo assim. A utilização mais correta do termo autotrófico é que eles seriam organismos independentes de material orgânico, que assimilam compostos chamados C-1 (CO2, CH4 ou metanol) como fonte de carbono. Parece que eu esqueci de colocar algum termo, não é? Onde entraria a energia luminosa nesta classificação mais ampla? Ela não entraria. Isto porque nem todos os organismos autotróficos utilizam a energia luminosa como fonte de energia. A fonte de energia pode vir da oxidação de compostos inorgânicos, como sulfeto, metano e hidrogênio.
Podemos dividir os organismos autotróficos em dois grandes grupos: os que dependem da energia luminosa (fotoautotróficos) e os que oxidam compostos inorgânicos e não dependem da energia luminosa (quimioautotróficos). Devido aos organismos quimioautotróficos, nem toda a vida de nosso planeta depende da energia do sol. A importância dos organismos quimioautotróficos na sustentação da cadeia alimentar foi primeiramente mensurada há mais de 30 anos em ecossistemas bem peculiares, as fontes hidrotermais. Elas se localizam em regiões de oceano profundo e se organizam em grandes comunidades de organismos, sendo considerada como uma das mais produtivas do mundo. Desta forma, toda esta biomassa, que inclui espécies de camarão, vermes tubulares, mexilhões é sustentada na ausência completa de luz.

Comunidade associada a uma fonte hidrotermal. Crédito: Nature
Após a descoberta das fontes hidrotermais, a importância da quimiossíntese foi estudada em outros ecossistemas, como sedimentos de ambientes aquáticos costeiros e corais. Como estes ambientes apresentam uma grande distribuição em todo o mundo, o estudo da real importância da quimiossíntese em termos globais tem sido o tema de muitos artigos em revistas de renome nos últimos anos. Ambientes de águas anóxicas como lagos profundos ou com grande taxa de decomposição têm altas taxa de metanogênese (produção de metano) por archeas, sendo ecossistemas onde a quimiossíntese poderia ter grande importância para a cadeia alimentar. O metano gerado por archeas metanogênicas é oxidado por bactérias metanotróficas, sendo uma fonte extra de energia e carbono para o ambiente. Este fato já foi comprovado por análises isotópicas, que conseguem determinar a origem do carbono. Quando o carbono (metano) é proveniente da metanogênese, ele tem um sinal baixo. Desta forma, organismos que apresentarem um sinal parecido, provavelmente tem uma dieta baseada em carbono com origem no processo metanogênico. Organismos com um sinal mais alto tem uma dieta mais direcionada para outras fontes de carbono, como fotossíntese. Um estudo publicado em 2008 na Ecology analisou o sinal das larvas de quironomídeos (Diptera) em 87 lagos e viu que as bactérias metanotróficas foram as maiores responsáveis pelo crescimento de biomassa das larvas.
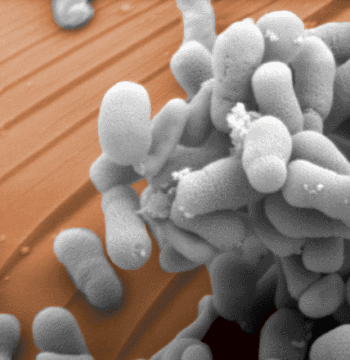
Bactérias metanotróficas (Methylosinus trichosporium OB3b). Crédito: Ezra Kulczycki
Acharemos o sinal de que o carbono em peixes e no homem pode ter origem bacteriana? Tudo indica que sim. Talvez em alguns anos descobriremos que organismos muitas vezes desconsiderados por nós seres “superiores” podem ser uma importante fonte de energia e carbono para a vida em nosso planeta, até mesmo para grandes vertebrados.
Para ler outros posts desta blogagem coletiva sobre a luz, clique aqui.
Referências:
Bastviken, D., Ejlertsson, J., Sundh, I., & Tranvik, L. (2003). METHANE AS A SOURCE OF CARBON AND ENERGY FOR LAKE PELAGIC FOOD WEBS Ecology, 84 (4), 969-981 DOI: 10.1890/0012-9658(2003)084[0969:MAASOC]2.0.CO;2
Dubilier, N., Bergin, C., & Lott, C. (2008). Symbiotic diversity in marine animals: the art of harnessing chemosynthesis Nature Reviews Microbiology, 6 (10), 725-740 DOI: 10.1038/nrmicro1992
Fenchel, G. M. and B. T.H. 1998. Bacterial biogeochemistry. The ecophysiology of mineral cycling. Academic Press.
Jones, R., Carter, C., Kelly, A., Ward, S., Kelly, D., & Grey, J. (2008). WIDESPREAD CONTRIBUTION OF METHANE-CYCLE BACTERIA TO THE DIETS OF LAKE PROFUNDAL CHIRONOMID LARVAE Ecology, 89 (3), 857-864 DOI: 10.1890/06-2010.1
muito legal me ajudou no trabalho de escola
Luiz, tenho que dizer que já passei por este teu texto em diversas situações de elaboração de materiais didáticos e cá estou eu novamente pra isso! Estou preparando algumas fichas de lição de casa pros alunos para o próximo ano e claro que vim parar aqui novamente 🙂
Parabéns pela síntese e didatismo! Sem dúvida os alunos curtirão muito ler mais este post (e serão aqueles mesmos a quem você gentilmente concedeu aquelas longas entrevistas!).
Beijão,
Tati
Hum… Entendi. O problema era que eu não sabia que processos biológicos também alteravam a razão isotópica natural… Obrigada pela explicação!
Olá liliane,
É exatamente isso! Não entendi porque ficou obscuro. Vou tentar exlicar de outra forma, voltando um pouco mais.
Tanto o carbono de massa 12 como o de massa 13 são estáveis. A proporção deles na natureza é razoavelmente constante (razão isotópica natural). Quase 99% de todo o carbono na natureza está na forma de carbono 12. O xis da questão está em como os processos biológicos alteram esta razão isotópica natural, através do fracionamento do carbono. Por exemplo, a forma que uma planta C3 e uma planta C4 fracionam o carbono altera o seu sinal, tendo como resultado uma razão isotópica diferente da natural. Assim, organismos que se alimentam mais de plantas C3 ou de C4, terão um sinal isotópico diferenciado ou uma razão entre o carbono 12 e 13 que compõem sua biomassa diferenciada.
Este fracionamento também ocorre com as archeas metanogênicas. Elas fracionam o carbono de uma forma diferente, resultando em uma razão isotópica diferenciada e distinguível dentre os outros processos. Neste caso, após o processo de metanogênese, a razão isotópica do carbono é desviada para mais carbono leve (carbono 12) do que carbono 13. Quando eu disse que a larva apresentava mais carbono 12 , quis dizer que, comparando com a razão isotópica natural, ela apresenta mais carbono 12, desta forma mais leve. O motivo seria a predominância em sua dieta de um carbono de origem nas archea metanogênicas, via bactérias metanotróficas.
E agora? 🙂
Oi Luiz, entendi o significado da expressão “sinal baixo”. No entanto, como vc chegou a conclusão, no final da sua resposta, de que a larva apresentará predominância de carbono 12, ainda está obscuro. Mas não se preocupe, eu sou Química, devem me estar faltando conhecimentos biológicos… rsrsrs
Olá Liliane,
Achei que não era necessário entrar em detalhes na metodologia de isótopos estáveis, mas vamos lá. Um sinal baixo indica um carbono “leve”, onde há o predomínio de Carbono 12 em detrimento de Carbono 13. A razão entre esses dois isótopos de carbono indicam a origem do mesmo. As archea metanogênicas fractionam o carbono durante o processo de metanogênese e este sinal é mantido ao longo da teia trófica. Desta forma, se uma larva de quironomídeo se alimentou de uma bactéria metanotrófica que anteriormente oxidou metano gerado por archeas metanogênicas, esta larva apresentará uma predominância de carbono leve (12).
Entendido?
🙂
Oi Luiz, adorei o post. Eu nunca tinha parado para pensar nesse detalhe (como nem todos os autótrofos dependem da luz). Só teve uma coisa que eu não entendi. Quando você disse “Quando o carbono (metano) é proveniente da metanogênese, ele tem um sinal baixo”, o que você quis dizer com “sinal”? Você se referiu à análise isotópica/abundância de carbono 13 ou RMN de carbono 13? Ou não tem nada a ver com isso?