


Fatores reguladores da virulência de Staphylococcus aureus
Antibióticos, bactérias, infecções, microrganismos são alguns dos assuntos que mais gosto, como meus leitores já devem ter percebido. Já foram postados vários textos neste blog sobre esta temática. Também já escrevi aqui sobre a bactéria patogênica Staphylococcus aureus, que pode ser extremamente nefasta, principalmente se a infecção por esta não for adequadamente tratada.S. aureus, como é conhecida, é uma bactéria danada. Ela possui vários mecanismos de virulência, como proteínas que atuam na adesão desta bactéria a superfícies e na invasão de tecidos de hospedeiros, exoproteínas que atuam contra mecanismos de imunidade, além de também possuir toxinas que causam hemólise (destruição dos hemócitos, ou glóbulos vermelhos) e que formam poros em membranas, permitindo o “vazamento” de íons e outras moléculas pequenas de células de hospedeiros. Para que cause infecção, é necessário que os fatores de virulência de S. aureus atuem de forma coordenada, até mesmo de maneira redundante, de tal forma que se um de seus fatores de virulência for afetado, os outros continuarão ativos.
Staphylococcus aureus
Descobriu-se que S. aureus tem um gene regulador de virulência, denominado agr (accessory gene regulator). Até pouco tempo atrás se sabia que a expressão do gene agr era regulada por um sinalizador químico. E se sabia de que tipo era este sinalizador: uma molécula pequena, da classe dos metabolitos secundários. Também se descobriu que, apesar do gene agr ter uma grande importância na regulação da virulência de S. aureus, este gene não é essencial para a expressão da virulência. São conhecidos isolados clínicos (de hospitais) de S. aureus desprovidos do gene agr.Recentemente, pesquisadores realizaram uma extensa análise do material genético (genoma) de S. aureus, de maneira a verificar a presença de genes reguladores da biossíntese (processo enzimático de várias etapas que é responsável pela formação das moléculas biológicas, como proteínas, açúcares, DNA, RNA, e também de metabolitos secundários) de peptídeos não-ribossômicos exclusivos de S. aureus. Analisando o genoma desta maneira, é possível se detectar a presença de genes que regulam a biossíntese de substâncias específicas, e propor a estrutura química das mesmas, uma vez que estes genes coordenam a ação de enzimas que atuam de maneira bem estabelecida. Foram analisados 50 genomas de S. aureus, e identificado um agregado de genes (gene cluster) responsável pela biossíntese de peptídeos não-ribossomais (ou seja, peptídeos que não são formados nos ribossomos, como são a grande maioria das proteínas, peptídeos muito grandes) ainda desconhecidos. O agregado de genes mostrou ter um gene de 7,17 Kb (1 Kb = 1000 bases nucléicas, que formam o DNA), que ocupa apenas 0,25% do genoma de S. aureus.
Este agregado de genes regula a formação de uma sintetase de peptídeos não-ribossomais (NRPS, non-ribossomal peptide synthetase) de 2389 aminoácidos. Esta NRPS mostrou ter sítios que atuam na formação de um dipeptídeo (um derivado de dois aminoácidos) com estrutura muito peculiar: é formado a partir dos aminoácidos valina e tirosina, que se condensam e dão origem a uma estrutura cíclica, de massa molecular de 262,17 u.m.a.(u.m.a. = unidade de massa atômica). Ou seja, a partir da análise do genoma de S. aureus, foi possível se encontrar genes que regulam a formação deste dipeptídeo, cuja estrutura pôde ser predita a partir dos resultados obtidos da análise do genoma.
Porém, era necessário que esta predição fosse confirmada.
Desta forma, os autores deste estudo cresceram S. aureus em meio de cultura, extraíram o meio de cultura com solventes orgânicos e analisaram os extratos do meio de cultura por cromatografia líquida (HPLC, high-performance liquid chromatography) acoplada a um detector de espectrometria de massas. Desta forma, para cada substância separada foi possível obter um espectro de massas, que indica a massa molecular da forma protonada das substâncias presentes no meio de cultura de S. aureus. Estas análises indicaram a presença de duas substâncias (dentre outras) no extrato orgânico do meio de cultura de S. aureus: uma de massa 262 e outra de massa 246. As duas mostraram ser muito parecidas pelo fato de serem pouco separadas na análise por HPLC, e também por apresentarem espectro no ultravioleta muito similar. Ou seja, talvez estas substâncias apresentassem estruturas muito parecidas, e tivessem uma biossíntese comum.
Esta hipótese foi confirmada após a obtenção das substâncias puras, uma separada da outra. Após isoladas, puderam ser analisadas por várias técnicas espectroscópicas, principalmente por diferentes experimentos de ressonância magnética nuclear (RMN). Estas análises indicaram que um dos dipeptídeos isolados era formado por tirosina e valina (aureusimina A), e o outro por fenilalanina e valina (aureusimina B). A diferença entre os dois é de apenas um átomo de oxigênio.
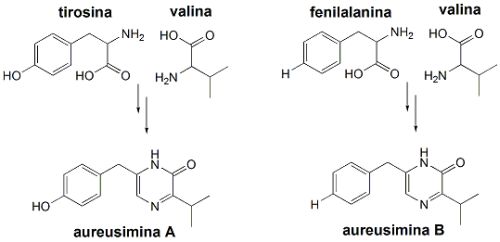
Para confirmar que estes dois dipeptídeos eram realmente biossintetizados por S. aureus, o gene ausA, supostamente responsável pela regulação da biossíntese destas duas substâncias, foi substituído por um gene artificial. E S. aureus não mais produziu as aureusiminas.Em seguida, foram feitos testes com as duas substâncias (aureusiminas A e B) para verificar como estas influenciavam na expressão do gene agr, que controla a virulência de S. aureus. Observou-se que a substituição do gene ausA de S. aureus, responsável pela biossíntese das auresiminas, por um gene artificial alterou completamente o padrão de expressão do gene agr, responsável pela virulência. Observou-se diminuição na formação de proteínas imunomodulatórias, de proteínas de adesão, bem como de proteínas líticas (que promovem o rompimento de membranas) e de citotoxinas (toxinas de células). Enquanto tais proteínas mostraram ter sua produção estimulada na presença das aureusiminas, na ausência destas substâncias as proteínas virulentas mostraram ser produzidas em quantidades muito menores.
Claramente as aureusiminas mostraram atuar na regulação das proteínas participantes do processo de virulência de S. aureus.
Normalmente, S. aureus “original”, sem ter seu genoma alterado, promove hemólise. Quando o gene ausA foi suprimido, a hemólise promovida por S. aureus sem este gene também foi suprimida. Quando se adicinou as aureusiminas no meio de crescimento de S. aureus com seu genoma alterado (com o gene ausA deletado), voltou-se a observar a hemólise. Claramente as aureusiminas participam diretamente da ativação dos processos de virulência de S. aureus.
E mais: quando camundongos sadios foram infectados com S. aureus “original” (sem ter seu genoma alterado), os bichinhos apresentaram grande quantidade de unidades formadoras de colônia (um indicativo de proliferação bacteriana) em seus rins, fígado, baço e coração. Quando os camundongos foram infectados com S. aureus mutada, com o gene ausA suprimido, observou-se que os rins apresentaram grau de infecção, mas os outros três órgãos muito menos, principalmente o coração (praticamente sem infecção). Ou seja, a inibição da biossíntese das aureusiminas levou a um grau de infecção virulenta muito menor.
A descoberta que as aureusiminas controlam a virulência de S. aureus abre um enorme conjunto de possibilidades para o desenvolvimento de novas formas de tratamento de infecções por esta bactéria. Por exemplo, a eventual descoberta de inibidores da biossíntese das aureusiminas pode significar a abolição do uso de antibióticos para o tratamento de infecções por S. aureus. Embora tal perspectiva possa parecer distante, tudo depende do esforço de pesquisadores acadêmicos e de indústrias farmacêuticas sérias. O mecanismo de ação de tais agentes inibidores da virulência de S. aureus seria completamente diferente do mecanismo de ação dos antibióticos. Isso porque os inibidores da biossíntese das aureusiminas interferem no processo bioquímico de formação destas substâncias, muito mais difícil de sofrer uma mutação benéfica para continuar promovendo a virulência. Desta forma, não seria mais necessário utilizar antibióticos, dos quais as bactérias adquirem resistência (através de mutações) em poucas gerações.
Infecção causada por S. aureus (nojenta!)
Embora o trabalho realizado com S. aureus pareça muito complicado, realmente não é. As atuais ferramentas bioquímicas permitem a realização de um trabalho deste em tempo relativamente curto. Na verdade, os autores mencionam um trabalho publicado em 2008 como fonte das informações preliminares nas quais se basearam para realizar seu estudo. A referência original indica que este trabalho (Novick e Geisinger) foi publicado on-line em agosto de 2008. Ou seja, o estudo realizado com S. aureus levou, no máximo, menos de dois anos para ser desenvolvido. Uma boa idéia e abordagens e estratégias de trabalho bem delineadas levaram à publicação de um artigo extramemente relevante na Science, uma vez que as infecções causadas por S. aureus são consideradas um dos maiores problemas de saúde pública no mundo.O que certamente fez a diferença para um trabalho deste ter sido feito em tão pouco tempo? Acesso a instrumentação e aos materiais (reagentes, linhagens bacterianas, camundongos) necessários para o desenvolvimento deste projeto. Sem entraves, sem burocracia, sem demora, sem falta de dinheiro.
 Wyatt, M., Wang, W., Roux, C., Beasley, F., Heinrichs, D., Dunman, P., & Magarvey, N. (2010). Staphylococcus aureus Nonribosomal Peptide Secondary Metabolites Regulate Virulence Science, 329 (5989), 294-296 DOI: 10.1126/science.1188888
Wyatt, M., Wang, W., Roux, C., Beasley, F., Heinrichs, D., Dunman, P., & Magarvey, N. (2010). Staphylococcus aureus Nonribosomal Peptide Secondary Metabolites Regulate Virulence Science, 329 (5989), 294-296 DOI: 10.1126/science.1188888
Novick, R., & Geisinger, E. (2008). Quorum Sensing in Staphylococci Annual Review of Genetics, 42 (1), 541-564 DOI: 10.1146/annurev.genet.42.110807.091640
A química supramolecular das aranhas
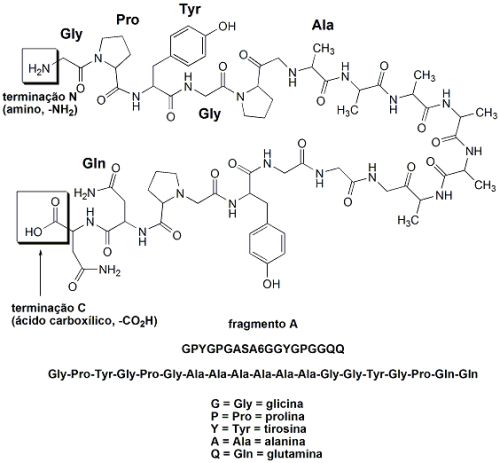
O fio das teias de aranhas é um material absolutamente excepcional: tem uma resistência mecânica proporcionalmente maior do que a do aço quando se leva em conta sua baixa densidade. Um único fio de teia de aranha alinhado sobre a linha do equador teria a massa de apenas 500 gramas. Os fios da teia de aranhas são feitos de proteínas: uma cadeia polipeptídica que consiste de uma sequência repetida de dois fragmentos peptídicos, juntos chamados de “sequência AQ”. O fragmento A é hidrofóbico (hidro=água; fobia=aversão. Ou seja, aversão à água, um fragmento que não admite moléculas de água associadas), que apresenta muitas unidades do aminoácido alanina (A): GPYGPGASA6GGYGPGGQQ (cada letra corresponde a um aminoácido). O fragmento Q é hidrofílico (hidro=água; filo=afinidade. Ou seja, com afinidade por água, é um fragmento que apresenta moléculas de água associadas através de ligações de pontes de hidrogênio), rico em glutamina e em glicina: (GPGQQ)4. Esta estrutura AQ é a “parte central” da fibra do fio da teia de aranha, e sua estrutura e função lembram a do colágeno (proteína espalhada pelo corpo dos humanos que forma o tecido conjuntivo). A fibra dos fios da teia de aranha são formados por 12 fragmentos AQ, os quais terminam com fragmentos diferentes, que atuam como “sinais químicos” e partes diferentes constituídas por grupos amina livres (-NH2) e grupos carboxila lvres (-CO2H). Estas partes diferentes formadas pelos grupos amina e carboxila livres participam de funções diferentes, como controle da solubilidade da proteína e da formação da fibra que dá origem ao fio da teia de aranha.
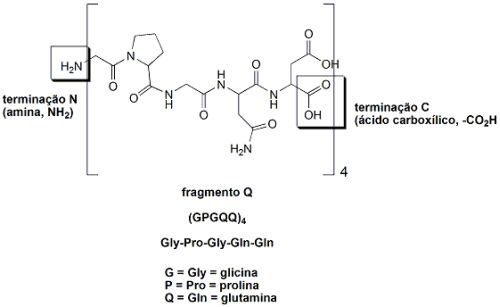
Por exemplo, a estrutura da terminação C (ácido carboxílico, -CO2H) não repetida (NR-C) da fibroína da aranha Aaraneus diadematus foi estudada em solução por ressonância magnética nuclear (RMN). A técnica de RMN detecta os núcleos de alguns átomos (mas não de todos), como por exemplo os de hidrogênio (1H). É possível se “medir” a forma como átomos de hidrogênio de uma molécula orgânica interagem entre si, e com isso entender como esta molécula orgânica se comporta em solução (por exemplo, em uma solução salina). Desta forma, é possível se verificar como partes das proteínas das aranhas “se dobram” em solução (foi o que foi feito). Foi observado que dois NR-C da fibroína de A. diadematus formam uma estrutura dimérica, altamente simétrica, com um formato de barril, constituído por duas hélices da proteína, unidas entre si por uma ponte dissulfeto (pontes dissulfeto, S-S, são ligações enxofre-enxofre formadas entre dois aminoácidos cisteína. Estas pontes dissulfeto aumentam a rigidez das proteínas). Este “formato de barril” da estrutura das proteínas das fibras dos fios da teia fazem com que as partes hidrofóbicas (A) fiquem agregadas, enquanto que a parte hidrofílica (Q) fica fora do barril, de maneira a que possa se agregar a moléculas de água.
Para que o fio da teia seja formado, é necessário que o pH do ambiente de formação seja muito bem regulado. Foram feitos experimentos com mini-spindroínas (proteínas bem menores do que as spindroínas, que formam os fios), que mostraram que quando a terminação N (amino, -NH2) está participando da formação da proteína, o pH do ambiente deve ser 6,0 (levemente ácido).De outra forma, em pHs mais baixos (mais ácidos) ou maiores (mais básicos), a proteína não se forma. Todavia, estes efeitos são reversíveis. Ou seja, se por um acaso o pH do ambiente se altera, e a formação da proteína é interrompida, a formação da proteína continua quando o pH do ambiente volta a ser 6,0. Já a terminação C (ácido carboxílico, -CO2H) participa da extensão da proteína, ou seja, do aumento do tamanho desta.
A formação das proteínas do fio da teia ocorre dentro de glândulas das aranhas, e são estocadas como oligômeros (pequenos polímeros) de alta densidade, com uma estrutura micelar (uma micela é uma estrutura esférica supra-molecular, ou seja, que faz uso de muitas moléculas de um determinado tipo, chamadas de detergentes, por terem uma extremidade carregada positiva ou negativamente, hidrofílica, e uma “cauda” sem carga, hidrofóbica), formando uma microemulsão. Cada micela contém várias proteínas dos fios da teia, e as micelas são estabilizadas pela presença dos grupos N (amina, -NH2) e C (ácido carboxílico, -CO2H) terminais. A auto-agregação das proteínas para formar as micelas é reversível.
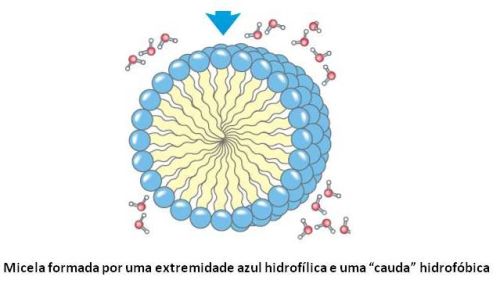
Dentro das aranhas, a formação das fibras de proteínas não é apenas determinada pelas mudanças químicas do ambiente, mas também por estímulos mecânicos em um tipo de sinergia (ação conjunta de forças) de efeitos macroscópicos e microscópicos, ilustrados na figura a seguir. Para entender as propriedades formadoras de fibrilas da cadeia de proteínas, é preciso compreender como a extrusão do complexo micelar supra-molecular acontece: estresse de cisalhamento (cisalhamento é um movimento de “escorregar para frente e para trás” ao longo do comprimento de um eixo), extrusão de água e troca de íons estimulam a formação das fibras dos fios da teia. A terminação C (ácido carboxílico) faz com que a cadeia AQ não mude de viscosidade durante o processo de cisalhamento. Por outro lado, a cadeia protéica já formada dá origem a uma forma globular que leva ao surgimento de agregados fibrosos quando o terminal C (ácido carboxílico) está presente.
As aranhas formam uma proteína extremamente específica para dar origem às fibras dos fios da teia. Estas proteínas são o resultado da fusão de um bloco de um co-polímero (AQ) anfifílico (anfi=ambos; filo=afinidade. Ou seja, um polímero que é ao mesmo tempo hidrofílico e hidrofóbico) e dois “barris” diméricos, com proteínas dobradas na forma de alfa-hélices. Nas glândulas da aranha, as proteínas formam um conjunto de uma microemulsão supramolecular. A aranha então aplica fortes forças de cisalhamento ao mesmo tempo que realiza a extrusão da proteína de suas glândulas. Esta extrusão é acompanhada da expulsão de água da microemulsão, levando ao desdobramento das estruturas de barris, fazendo com que os fragmentos hidrofílicos se projetem de dentro para fora. Ocorre então uma mudança macroscópica na forma da emulsão, levando a uma polimerização específica que dá origem aos fios da teia com sua enorme resistência mecânica.

A transição na forma das proteínas dos fios da teia, a partir de um complexo supramolecular solúvel em água na forma de micelas para formar fibras longas, mostra como estas proteínas podem mudar de forma de acordo com mudanças no ambiente químico e sob forças mecânicas. Além disso, o estudo realizado mostrou não somente as particularidades de como os fios das teias de aranha e como a teia é formada, mas também como as aranhas trabalham com os fios para formar a teia mais eficiente para capturar suas presas. Embora a formação de fibrilas protéicas sempre foi vista co
mo uma disfunção estrutural, levando à perda de forma tridimensional das proteínas que as formam, as aranhas se utilizam de tais propriedades para formar os fios de suas teias, altamente resistentes.
Perguntas que ainda restam para serem respondidas sobre as teias de aranha incluem: será que as terminações N (amina) e C (ácido carboxílico) operam de maneira cooperativa, ou reagem diferentemente sob um determinado estímulo químico? Como a agregação supramolecular das cadeias protéicas individuais garante a formação de uma estrutura em rede para dar origem a fios de até 1 metro de comprimento? Como que um fio formado de maneira tão específica pode responder tão bem a diferentes presas capturadas pelas aranhas? A qualidade dos fios das teias depende de como esta é formada pela aranha e fatores ambientais, bem como da alimentação das aranhas. Mas a qualidade intrínseca dos fios é determinada pela sua composição de aminoácidos. A compreensão de como os componentes dos fios da teia pode dar origem ao processo de formação da teia, com suas propriedades mecânicas, ainda permanece um enigma para as ciências dos biomateriais.
Só o homem-aranha conhece estes segredos.
Silvers, R., Buhr, F., & Schwalbe, H. (2010). The Molecular Mechanism of Spider-Silk Formation Angewandte Chemie International Edition DOI: 10.1002/anie.201003033
Quanta vida na Terra?
Mas May também gosta de apreciar as brincadeiras alheias com números deste tipo. E de fornecer dados sem citar referências. Em seu mais recente artigo, “Tropical Arthropod Species, More or Less?”, May inicia suas divagações sobre quais impressões teriam extra-terrestres ao chegar na Terra. Segundo ele (May), estes teriam como primeira curiosidade saber quantas formas distintas de vida existem na Terra. Embora eu não concorde com este questionamento de May (acho que extra-terrestres teriam mais curiosidade em saber que TIPOS de vida existem na Terra), entendo a introdução de seu artigo como uma provocação sobre a nossa (humanos, terráqueos) própria ignorância sobre este assunto. Afinal, ao afirmar que “um terço dos taxonomistas trabalham com vertebrados (1% de todas as espécies biológicas), outro terço dos taxonomistas com plantas (cerca de 10% de todas as espécies biológicas), e o último terço com invertebrados (cerca de 90% de todas as espécies)”, May está querendo dizer que há pouca gente para conhecer tanta diversidade biológica. May acredita que o número total de espécies biológicas classificadas seja de 1,6 a 1,7 milhões, com cerca de 15.000 a mais a cada ano. Ao se levar em conta um número (bem conservador) de 3 milhões de espécies vivas, no total, estamos longe de conhecer a diversidade biológica da Terra.
May se detém na análise realizada por Hamilton e seus colaboradores, que se utilizou de estimativas e abordagens estatísticas modernas para estimar o número de artrópodes da Terra. Segundo a Wikipédia,
Os Artrópodes (do grego arthros: articulado e podos: pés, patas, apêndices) são animais invertebrados caracterizados por possuírem membros rígidos e articulados. São o maior grupo de animais existentes, representados pelos gafanhotos (insetos), aranhas (arachnida), caranguejos (crustáceos), centopeias (quilópodes) e embuás (diplópodes), somam mais de um milhão de espécies descritas (apenas mais de 890.000 segundo outros autores). Mais de 4/5 das espécies existentes são Artrópodes que vão desde as formas microscópicas de plâncton com menos de 1/4 de milímetro, até crustáceos com mais de 3 metros de espessura.

Hamilton e seu time partiram das idéias de Erwin, que tentou estimar o número de espécies de besouros (o maior grupo dos artrópodes) depois de realizar um estudo da distribuição vertical destes animais sobre uma única espécie de árvore tropical. Erwin estimou o número de espécies de artrópodes tropicais entre 30 e 100 milhões. A escolha de Erwin baseou-se no fato que a diversidade de artrópodes, em particular de besouros, nas regiões tropicais é muito maior do que em outras regiões da Terra. Já o time de Hamilton realizou vários estudos sobre a distribuição de besouros e outros artrópodes em árvores de diferentes países, como a Nova Guiné (na África; ou seria Papua Nova Guiné, no Pacífico Sul, perto da Indonésia?), Brasil, Panamá, Venezuela, Sulawesi, entre outros países.
Erwin estimou que 40% dos artrópodes seriam besouros. Os estudos do time de Hamilton levaram à conclusão, utilizando técnicas modernas e sofisticadas de análise estatística, que o número de besouros corresponderia de 22 a 60% dos artrópodes. Na média, muito próximo do resultado de Erwin. Este estimou que o número aproximado de árvores tropicais seria de 50.000. Hamilton e seus colaboradores chegaram a uma estimativa entre 43.000 e 50.000. A principal diferença nos resultados dos dois estudos, publicados com um intervalo de 28 anos, é que os besouros parecem ser muito menos especializados (segundo Hamilton) do que pareciam ser (segundo Erwin). Ou seja, besouros podem se alimentar em várias árvores de espécies diferentes, e não sempre na mesma e única espécie de árvore. Desta forma, o número de espécies de artrópodes estimado pelo grupo de Hamilton é bem menor do que entre 30 e 100 milhões (estimado por Erwin). Segundo Hamilton, este número estaria próximo de 3,7 milhões de espécies, com 90% de probabilidade de estar entre 2,0 e 74 milhões de espécies, quando se utilizou de um primeiro modelo de análise. Para um segundo modelo de análise, o número foi estimado de 2,5 milhões de espécies de artrópodes, com 90% de probabilidade de se situar entre 1,1 e 5,4 milhões de espécies (de artrópodes).
De acordo com os resultados apresentados por Hamilton (e discutidos por May), cerca de 2/3 das espécies de artrópodes da Terra ainda não foram descritas. Segundo May, este número seria um reflexo da distribuição desigual de taxonomistas para a descrição dos diferentes grupos de organismos vivos. Mas por outro lado, também que as agências financiadoras consideram taxonomia uma “ciência menor, apenas descritiva, e pouco geradora de conhecimento associado”. Por isso que a taxonomia é uma ciência em vias de extinção. E, em paralelo, as espécies biológicas também, mas não sabemos ao certo quantas nem quais.
May, R. (2010). Tropical Arthropod Species, More or Less? Science, 329 (5987), 41-42 DOI: 10.1126/science.1191058
