


Vendo a vida como ela é
Quando a subjetividade interfere na análise de dados e fenômenos científicos, a apreciação dos mesmos pode ficar comprometida devido a uma interpretação enviesada. O entendimento dos processos biológicos, em particular do funcionamento das células, durante um bom tempo sofreu a interferência da interpretação através da teoria do vitalismo. Embora esteja bem estabelecido o conhecimento atual sobre a célula de que esta é constituída de um pequeno volume de líquido contendo milhares de moléculas orgânicas e inorgânicas, que troca matéria e energia com seu ambiente, esta noção é bastante recente. Segundo Marc Fontecave (Université Joseph Fourier, Grenoble, França) ao se discutir sobre o conhecimento da vida em um contexto histórico, é possível se abordar este tema do ponto de vista vitalista e do ponto de vista reducionista.
Atualmente se considera que as estruturas e os processos químicos explicam muito bem a vida no seu âmago. Isso porque a química tem a capacidade de explicar tanto a “matéria com informação” como a “matéria de transformação”. Nesta última pode-se encontrar não somente moléculas orgânicas como também inorgânicas oriundas dos processos de transformação; no primeiro caso, encontram-se as moléculas envolvidas na manutenção da vida. Tal “reducionismo” é inerente à abordagem científica, quando se objetiva compreender sistemas tão complexos como são as células.
Os vitalistas defenderam, durante um bom tempo, a existência de uma “força vital” que seria a força motriz que impregnaria todos os seres e processos vivos. Por exemplo, o fisiologista francês Marie François Xavier Bichat (1771-1802) afirmou, no fim do século XVIII, que “A ciência dos corpos organizados não deve ser considerada da mesma forma que a ciência dos corpos inorgânicos (..) Fenômenos físicos e químicos obedecem às mesmas leis, mas existe uma grande lacuna entre estas leis e as leis da vida”. Por outro lado, pesquisadores como Justus Von Leibig (1803-1873), Marcelin Berthelot (1827-1907) e Claude Bernard (1813-1878) acreditavam que os processos vivos podiam ser entendidos apenas com base nas leis físicas e químicas.

A história da química biológica é marcada pela disputa entre vitalistas e reducionistas. Até quase o final do século XVIII, a distinção entre matéria viva e inanimada era desprovida de uma fronteira definida. Isso porque as ferramentas de investigação científica ainda eram muito limitadas. Ao mesmo tempo, as ciências mecânicas descobertas por Newton, da Vinci e outros, não serviam para descrever de maneira apropriada os sistemas vivos. Assim, postulava-se a existência de uma força espiritual que impregnava todas as formas de vida. O trabalho de pesquisadores químicos no final do século XVIII e no início do século XIX começou a modificar este quadro. Por exemplo, em 1783 Scheele (1742-1786) descobriu o ácido cítrico e a glicerina trabalhando com seu mentor, Bergman (1735-1784). Tais compostos foram denominados como sendo “orgânicos” para distingui-los dos inorgânicos, aqueles desprovidos de átomos de carbono. A síntese em laboratório dos compostos orgânicos era considerada como sendo impossível, pois era absolutamente necessária a atuação de uma força vital para a sua formação.
Porém, em 1828 Friedrich Wöhler (1800-1882), ao tentar produzir isocianato de amônia, aqueceu uma mistura de isocianato de prata e cloreto de amônia, obtendo uréia. A uréia já era conhecida desde 1783, quando foi pela primeira vez isolada a partir da urina animal. Foi a primeira vez que uma molécula orgânica havia sido preparada em laboratório, sem a “ajuda de uma força vital”. Em 1845 Herman Kolbe conseguiu sintetizar ácido acético a partir de carvão, hidrogênio e água na presença de sulfeto de ferro e cloro. Subsequentemente, Berthelot conseguiu sintetizar acetileno utilizando carbono e hidrogênio sob uma corrente elétrica. Os resultados obtidos por Wöhler, Kolbe e Berthelot foram o ponto de partida para o nascimento da química dos seres vivos, denominada de química biológica. Paralelamente, os pesquisadores alemães Schwann (1810-1882) e Schleiden (1804) estabeleceram a assim chamada “teoria celular”, de que todo e qualquer organismo vivo é constituído por células.
Curiosamente, ainda um bom tempo depois, Louis Pasteur (1822-1895) reavivou a teoria vitalista, ao utilizar seu preceitos para “explicar” a dissimetria molecular intrínseca às substâncias quirais, bem como para explicar os processos de fermentação microbiológica.
O conceito vitalista passou a ser fortemente questionado, e por fim abandonado, quando foram descobertas as enzimas e o DNA. Eduard Buchner (1860-19170, prêmio Nobel em 1917) descobriu que extratos de leveduras livres de células mantinham sua capacidade de converter glicose em álcool e dióxido de carbono, desqualificando a teoria vitalista de fermentação postulada por Pasteur. Mais tarde, em 1953, Watson e Crick descreveram a estrutura do DNA, que explica, a nível molecular, sua capacidade de replicação, levando à geração de células de gerações descendentes contendo o mesmo material genético que suas células predecessoras. A estrutura do DNA estabelecida por Watson e Crick explica suas propriedades físicas e químicas, bem como sua capacidade de replicação, mutabilidade e sua capacidade de expressar a informação genética.
Embora reducionista, a abordagem científica que hoje descreve os processos celulares passou a ser mais recentemente considerada de uma nova maneira, holística, mais completa – através de uma biologia de integração, denominada de “biologia de sistemas”. A biologia de sistemas explica que os organismos vivos são uma unidade na sua essência, e, apesar de poderem ser completamente descritos em termos de todos os seus componentes químicos e bioquímicos, tais componentes não explicam, per se, o funcionamento dos sistemas vivos. É necessário conhecer como estes componentes se relacionam de múltiplas formas, de maneira a compreender os processos biológicos – objetivo desta nova ciência, a biologia de sistemas, na qual toma parta a biologia química (que NÃO É o mesmo que química biológica).
Mesmo após ter sido cientificamente abandonado, o vitalismo ainda é adotado por correntes de pensamento político e anti-científico, muitas vezes de cunho religioso fundamentalista, e também por grupos que defendem a natureza de maneira extrema, dizendo que “compostos naturais não são químicos”. Tais ideologias podem ser facilmente conduzidas ao ostracismo se cientistas e educadores sérios trabalhassem de forma conjunta para prover uma melhor educação e conhecimento sobre os processos vivos para as pessoas em geral.
A descoberta que o genoma humano contém apenas 30.000 genes exemplifica muito bem o estado atual do conhecimento científico, uma vez que tal número não é aceito por diversas correntes ideológicas como sendo responsável para explicar a natureza humana. Todavia, a explicação científica para tal fato é que a complexidade dos organismos vivos resulta em mais do que a soma de seus componentes moleculares. Estes se relacionam através de processos emergentes e restritivos, através dos quais
surge a informação biológica e suas manifestações, que resultam na multitude de formas de vida que conhecemos hoje no planeta Terra.
 Fontecave, M. (2010). Understanding Life as Molecules: Reductionism Versus Vitalism Angewandte Chemie International Edition DOI: 10.1002/anie.200906869
Fontecave, M. (2010). Understanding Life as Molecules: Reductionism Versus Vitalism Angewandte Chemie International Edition DOI: 10.1002/anie.200906869
A ancestralidade única e comum de todos os seres vivos
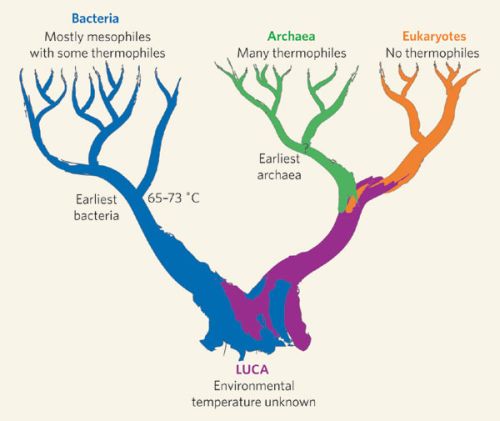
Apesar do conceito de UCA ter sido proposto há 150 anos, é amplamente aceito, e é uma das premissas mais importantes da TE. Além de ter sido corroborado inúmeras vezes, se fundamenta também por diversas constatações: (1) pela concordância entre filogenia e biogeografia; (2) bem como entre filogenia e os achados paleontológicos; (3) pelos achados de inúmeros fósseis que foram previstos que seriam encontrados; (4) pela classificação hierárquica de caracteres morfológicos; (5) pelas notáveis similaridades de estruturas biológicas que apresentam diferentes funções (homologias), e; (6) pela congruência constatada entre filogenia morfológica e filogenia molecular.
O conceito de UCA é ainda aceito pelo fato da constituição bioquímica dos seres vivos ser, na sua essência, a mesma: os mesmos açúcares, os mesmos aminoácidos, vários lipídios comuns, bem como seus “polímeros” correspondentes: proteínas, RNA e DNA. A “base bioquímica” da vida é uma forte evidência que esta se desenvolveu a partir de uma ancestralidade comum que deu origem a todos os seres vivos.
Muitas proteínas de organismos completamente diferentes apresentam alto grau de similaridade, a assim chamada homologia protéica (Osvaldo A. S. Filho e Ricardo B. Alencastro explicam o conceito em artigo publicado em português na revista Química Nova). Na homologia protéica, a similaridade na sequência de aminoácidos é verificada experimentalmente. Uma significativa similaridade na sequência de aminoácidos de proteínas de diferentes organismos vivos pode surgir não somente pela ancestralidade comum, mas também devido à convergência evolutiva associada aos processos de seleção natural, a restrições estruturais que acabam por influenciar a identidade da sequência de aminoácidos, bem como determinadas tendências nas mutações e até mesmo o acaso.
Até mesmo céticos que duvidam da UCA têm que admitir que proteínas universalmente conservadas, presentes em quase todas as classes de seres vivos, apresentam sequências muito similares, homólogas (homologia no sentido se similaridade estrutural devido à fidelidade ao arquétipo a partir do qual tiveram origem). Levando-se em conta que é possível se quantificar o grau de similaridade, ou homologia, entre proteínas, também é possível, a partir de uma tal quantificação, verificar o grau de homologia entre proteínas considerando-se o conceito UCA ou uma ancestralidade múltipla.
Esta foi a idéia de Douglas L. Theobald (Departamento de Bioquímica, Brandeis University).
Utilizando a teoria de seleção de modelos (TSM, do inglês, model selection theory), Theobald testou o conceito UCA sem assumir que a similaridade de sequência de aminoácidos em proteínas indicava uma relação genealógica. A TSM provê métodos para identificar uma possível hipótese como sendo a que melhor explica determinados dados experimentais. Para tanto, deve-se levar em conta que, quando diferentes modelos científicos podem ser empregados para uma determinada análise, dois fatores opostos devem ser considerados: concordância com os dados experimentais e parcimônia. A concordância de um modelo de análise com dados experimentais pode ser arbitrariamente melhorado ao se aumentar o número de parâmetros a serem considerados. Porém, tal procedimento carrega um problema intrínseco, que é o de aumentar sobremaneira a complexidade da análise. Por isso a parcimônia: quanto mais simples a análise dos dados, melhor.
Os métodos de seleção de modelos levam ambos fatores em consideração, e fornecem meios de balanceá-los estatisticamente, de maneira a encontrar uma hipótese que explique os dados ao mesmo tempo da maneira mais acurada e mais precisa. O uso de tais métodos permite a quantificação das evidências contra e a favor de modelos competidores (ou seja, que podem explicar o mesmo conjunto de dados, cada um de uma maneira diferente). Theobald aplicou três métodos de seleção de modelos para testar o conceito de UCA (LLR, log likelihood ratio; AIC, Akaike information criterion, e; LBF, log Bayes factor).
Utilizando os três modelos, Theobald analisou se os três domínios da vida na Terra, Eukarya (os eucariontes, organismos vivos com organelas diferenciadas com membranas), Bacteria (bactérias em geral, que são procariontes, ou seja, não apresentam organelas celulares diferenciadas) e Archaea (micro-organismos diferentes das bactérias, pertencentes a um grupo totalmente distinto), para ver se estes são melhor descritos de maneira relacionada, com uma ancestralidade comum (UCA), ou se estes três domínios da vida surgiram de maneira independente e paralela.
Theobald utilizou as seguintes premissas para sua análise:
a) sequências de proteínas mudam com o passar do tempo, através da substituição de aminoácidos de maneira gradual, que pode ser descrita com uma matriz 20 x 20 (uma vez que nas proteínas são observados 20 aminoácidos essenciais) definida por certas frequências equilibradas de aminoácidos e uma matriz simétrica de trocas de aminoácidos;
b) que novos genes geneticamente relacionados são gerados por duplicação durante a especiação em bifurcações ou duplicação de genes, e;
c) que substituições de aminoácidos não estão relacionadas em linhagens de organismos diferentes nem em sítios protéicos específicos.
Os testes de seleção de modelos avaliam como tais premissas explicam os dados experimentais obtidos quando se postula que vários sub-conjuntos de grupos taxonômicos e proteínas apresentam uma ancestralidade comum.
O que é interessante é que o conceito UCA possibilita uma origem múltipla para a origem da vida. Assim, se a vida teve vários “inícios”, necessariamente devem ter ocorrido sérias restrições ao longo da história evolutiva para que somente os organismos vivos resultantes de um único grupo destas origens tenham sobrevivido. Os outros se extinguiram. Uma outra explicação seria que múltiplas populações, com origens separadas e independentes, adquiriram capacidade de compartilhar material genético de maneira convergente, e consequentemente se tornaram uma única espécie, antes de divergir novamente.
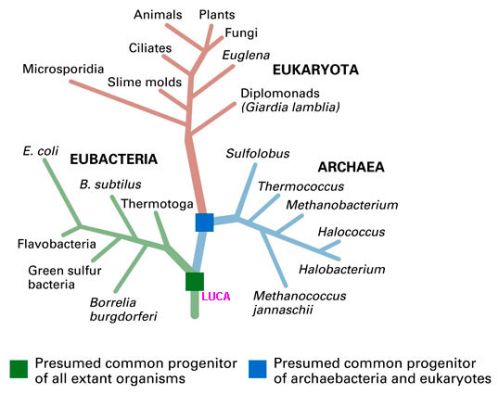
Todos os modelos examinados por Theobald são compatíveis com origens múltiplas e independentes, de acordo com qualquer uma das duas hipóteses. Os testes foram aplicados para se discriminar entre uma ancestralidade múltipla e uma ancestralidade comum universal (UCA).
Porém, não se analisou uma origem da vida única ou múltipla. Tampouco se considera a UCA como sendo de um único organismo, ou indivíduo. Na verdade, a ancestralidade comum universal se refere à de um grupo ancestral único. Ou a uma popula
Tomates verdes, tomates vermelhos

Tomates são ricos em carotenóides, tocoferóis e polifenóis. Os carotenóides são antioxidantes lipofílicos (solúveis em gordura), são precursores de substâncias aromáticas e exercem diversas funções bioquímicas nas plantas. Os tocoferóis também são antioxidantes lipofílicos. Já os polifenóis apresentam maior caráter hidrofílico (solúveis em água), principalmente quando ligados a moléculas de açúcares. Durante o processo de maturação, ocorrem diversas mudanças nos tomates que alteram sua composição de carotenóides, tocoferóis e polifenóis.
Em geral a concentração destas substâncias é maior em variedades selvagens (não cultivadas) de tomates. Observando variedades de tomates vermelhos, amarelos e verdes, verificou-se que as clorofilas a e b, bem como a feofitina, diminuem de concentração à medida que a variedade vermelha vai amadurecendo. Ao mesmo tempo, a concentração total de carotenos vai aumentando a partir do momento em que o amadurecimento se inicia até o seu ápice. Durante este processo, grandes quantidades de licopeno são sintetizadas e armazenadas na forma de cristais.
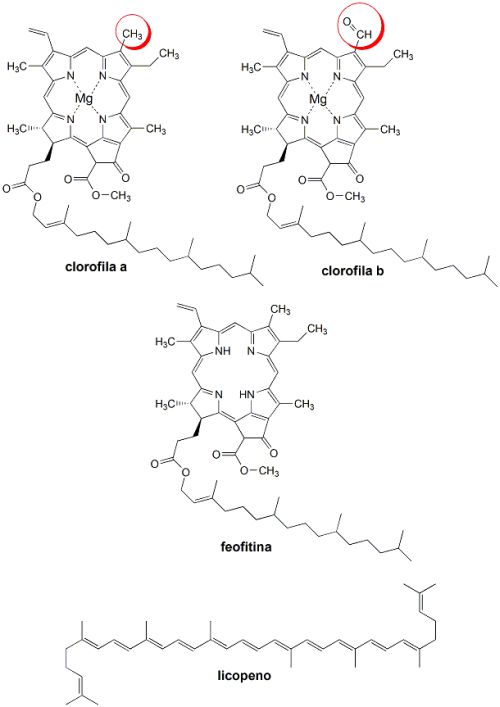
Já na variedade de tomate verde, tanto as clorofilas quanto a luteína como os carotenos diminuem de concentração durante o amadurecimento.
Tanto a espécie vermelha quanto a verde apresentam mudanças significativas a partir do início do amadurecimento. Ocorrem mudanças nas estruturas dos cloroplastos (organelas celulares responsáveis pela fotossíntese), e os genes responsáveis pela regulação da biossíntese de carotenos são ativados. Os tilacóides (sub-estruturas dos cloroplastos) incham, seus lipídios (gorduras) começam a se degradar, formando gotículas gordurosas com carotenóides livres ou ligados às gorduras. Este é um processo de senecência (envelhecimento) do tomate, que leva também a uma diminuição de sua quantidade de amido.
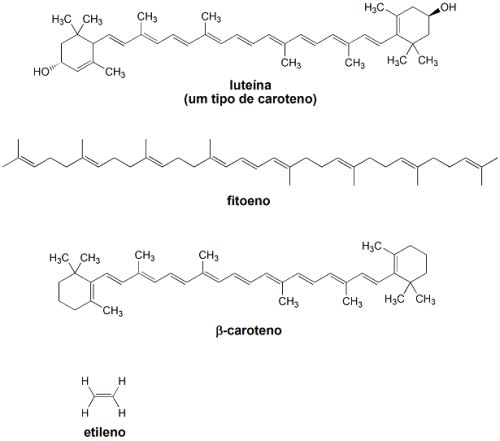
As mudanças que ocorrem no processo amadurecimento -> senecência levam a um acúmulo progressivo de licopeno, sendo que no tomate totalmente maduro a concentração de licopeno corresponde a 50% do total de carotenóides. Destes, a maior parte é de fitoeno, luteína e ß-caroteno. Como resultado da degradação do sistema de fotossíntese do tomate, ocorre uma mudança drástica na relação ß-caroteno/luteína: no início do amadurecimento esta relação é da ordem de 1,5, e no auge do amadurecimento esta relação se situa na faixa 0,37-0,69. Tais medidas podem ser úteis para se acompanhar, de maneira quantitativa, o processo de amadurecimento deste fruto.
No caso dos tomates verdes, estes não são desprovidos dos genes que regulam a formação de carotenóides. Porém, estes genes se encontram sub-expressos. Tal diferença também influencia no processo de maturação dos tomates verdes, que é fortemente influenciada pelo etileno, um hormônio vegetal. Em tomates verdes a produção de etileno é muito reduzida e, por isso, o fruto verde praticamente não amadurece.
À medida que o amadurecimento dos tomates acontece, ocorre uma diminuição significativa nos seus polifenóis.
Sumarizando, os frutos do tomateiro sofrem uma alteração bioquímica significativa durante seu amadurecimento, invertendo as quantidades de clorofila, tocoferóis, carotenóides e polifenóis. Quando ainda verdes, os tomates apresentam grandes quantidades de clorofilas, tocoferóis e polifenóis. Quando maduros, as clorofilas, tocoferóis e polifenóis praticamente desaparecem, restando grandes concentrações de carotenóides. Tomates vermelhos maduros apresentam significativa ação antioxidante, mas esta também se manifesta quando os frutos ainda estão verdes. Isso porque na fase verde a atividade antioxidante se deve aos polifenóis, mas na fase madura se deve aos carotenóides. No caso dos tomates verdes e amarelos, a atividade antioxidante se deve, sempre, à presença de polifenóis.
Tais mudanças influenciam diretamente a qualidade nutricional dos tomates e a maneira de consumi-los. Tomates verdes podem ser consumidos sem godura vegetal, como azeite de oliva. Já os tomates vermelhos devem ser consumidos com óleo vegetal, para um melhor aproveitamento de suas qualidades nutricionais.
Bon appetit!
Meléndez-Martínez, A., Fraser, P., & Bramley, P. (2010). Accumulation of health promoting phytochemicals in wild relatives of tomato and their contribution to in vitro antioxidant activity Phytochemistry DOI: 10.1016/j.phytochem.2010.03.021
Epicuro e o questionamento do “argumento do design”

A concepção filosófica de Sócrates foi elaborada para rebater a visão de mundo de Demócrito e Leucipo, fundadores do atomismo, segundo a qual o mundo material e a vida resultam da existência de partículas muito pequenas, os átomos (tomo = divisão; a-tomo = indivisível). Da mesma forma como Sócrates, Platão, aluno do primeiro, promoveu uma filosofia criacionista. Platão afirmou que a ordem se originava a partir do caos através da matéria, e estabeleceu a idéia de um Demiurgo (uma entidade sobrenatural, divina), que seria a maior de todas as causas e responsável pela formação do mundo.
Depois de Platão, Aristóteles propôs que tudo no mundo tinha um propósito. Esta é uma noção particularmente importante para entender a idéia de um projeto (design), que requer um projetista (designer). “Teleologia” é o termo cunhado para exprimir a idéia de uma razão final, um propósito, um objetivo, e que permeia os argumentos em favor de um “design”. Desta forma, o “design” seria a maneira através da qual um “designer” (projetista) manifesta seus propósitos, seus objetivos. Logo, qualquer argumento criacionista traz teleologia na sua essência. Aristóteles é considerado o pai da teleologia.
Os Estóicos foram aqueles que apresentaram o “argumento do design” da maneira mais explícita. Propuseram uma visão teleológica de um deus providencial, a existência do qual podia ser verificada através da “evidência do design” na natureza. Cícero, em sua obra “A natureza dos deuses”, defende abertamente o argumento em favor do design.

Por outro lado, Epicuro e seus seguidores, principalmente Lucrécio, desenvolveram uma lógica argumentativa extremamente sofisticada para questionar o argumento do design. A filosofia de Epicuro procurou apresentar argumentos de que os fenômenos naturais estavam livres do destino estabelecido pelos deuses e pelo determinismo mecanicista. Epicuro e Lucrécio rejeitaram qualquer explicação teleológica para o mundo material e os fenômenos da natureza. Epicuro procurou descrever o mundo em termos de seus processos naturais e não em termos de suas causas finais, uma vez que estas não podiam ser conhecidas, mas apenas ser objeto de especulação. Segundo Epicuro, uma explicação científica deveria ser obtida a partir da experiência, e estar de acordo com os princípios gerais que regem os fenômenos naturais, sem que se procurasse inferir quais seriam as possíveis causas (desconhecidas) destes fenômenos (naturais) ou a razão pela qual estes existiriam ou seriam do jeito que são.
Sem os conhecer de maneira deliberada, Epicuro criou os modernos conceitos de emergência e contingência no cerne de sua argumentação sobre o mundo natural. Segundo Epicuro, “nada permanece sempre da mesma forma, tudo está em mudança. Tudo é transformado pela natureza e é forçado a seguir novos caminhos”. Ainda segundo Epicuro, “a ação [dos fenômenos naturais] decorre como resultado da organização”, segundo a qual “a organização cada vez mais complexa de formas de vida superiores permite o surgimento [emergência] de novas maneira de vida, novas funções e comportamentos” (Foster, Clark, York, Critique of Intelligent Design, páginas 55-56).
Epicuro forneceu bases para explicar o desenvolvimento da sociedade humana, como o surgimento da linguagem, o altruísmo e a aquisição do fogo. Rejeitou toda interferência divina na história da humanidade, e promoveu a liberdade de pensamento, livre da superstição e da religião de estado. Foi o primeiro filósofo que propôs a noção de incerteza nos rumos da natureza, e também foi o primeiro a entender a contingência nos fenômenos naturais. Ou seja, que estes fenômenos resultam de uma série de fatores, coincidentes, que dão origem a um fenômeno ou a um processo particular.
Em seu ensaio “Materialismo e Revolução”, Jean-Paul Sartre afirma que Epicuro foi o primeiro filósofo que promoveu a idéia de libertação do homem de seus medos e aprisionamentos, e o primeiro que tentou abolir a escravidão (J. P. Sartre, Literary and Philosophical Essays, página 207). Sempre procurou combater uma religião do estado, como proposto por Platão em suas Leis. Segundo Epicuro, os deuses não tinham relação com o universo natural, mas somente com um mundo sobrenatural, que não poderia ser explicado, uma vez que a razão humana não têm acesso aos desígnios divinos. Assim, Epicuro não era ateu. Apenas defendia a idéia que o mundo subrenatural de deus não poderia ser alcançado pela razão.
Epicuro foi o primeiro pensador a introduzir o conceito de “contrato social”, e a afirmar que a moralidade é moldada historicamente e determinada pela prática social humana.

Ao rejeitar o “argumento do design”, Epicuro se estabeleceu como sendo o opositor aos Platonistas. Por isso, foi condenado pela Igreja dos Pais por sua rejeição à divina providência e à imortalidade da alma. Epicuro é considerado como sendo o maior crítico do “argumento do design” na idade antiga. Dante Alighieri (1265-1321), em seu clássico “Inferno”, descreve Epicuro e seus seguidores condenados à tortura eterna em caixões abertos.
A partir do surgimento do cristianismo, as idéias de Epicuro foram sendo deixadas de lado, devido à dominação da igreja. Durante a Idade Média as idéias de Epicuro foram praticamente esquecidas, até a redescoberta do manuscrito “De Rerum Natura” em 1417, de autoria de Lucrécio, principal seguidor de Epicuro. As idéias de Epicuro renasceram com o Iluminismo, e se tornaram a base para o desenvolvimento científico. Filósofos como Francis Bacon, Pierre Gassendi, René Descartes, Thomas Hobbes, Jean-Jacques Rousseau, David Hume e Immanuel Kant incorporaram muito da filosofia de Epicuro quando da fundação da filosofia da ciência tal como é atualmente conhecida. Assim, Epicuro é conhecido como sendo o pai das ciências naturais empíricas.
Bibliografia consultada
J. B. Foster, B. Clark, R. York, Critique of Intelligent Design, Monthly Review Press, New York, 2008.
J. P. Sartre, Literary and Philosophical Essays, New York, Criterion Boos, 1955.
A Natureza é Promíscua
Com o surgimento das ciências genômicas e a descoberta da reação de polimerização em cadeia, foi possível se explorar a codificação genética de inúmeros organismos vivos, e também de entender como funciona a expressão genética em geral (ou seja, qual é a função de um determinado gene, e qual o processo ou característica fenotípica que ele regula). A partir deste momento a biologia sofreu uma verdadeira revolução e o volume de informações sobre regulação gênica de processos biológicos aumentou exponencialmente.
Uma das grandes questões que surgiram após a descoberta do DNA foi: será que organismos vivos podem trocar DNA entre si? Ou seja, trocar genes? Aos poucos se verificou que esta hipótese era verdadeira. Bactérias de diferentes espécies frequentemente trocam genes entre si. Mas tarde se observou tal fato para fungos também. O surpreendente foi a observação de troca de genes entre fungos e bactérias, uma vez que são organismos que pertencem a Reinos diferentes (apenas para lembrar, a classificação dos organismos segue uma hierarquia: Reino, Filo, Classe, Ordem, Família, Gênero, Espécie). Ou seja, são organismos muito distantes do ponto de vista evolutivo. Mais recentemente, se observou a troca de genes entre bactérias e plantas. Esponjas marinhas, por exemplo, são animais. Mas esta definição é um tanto complicada, porque determinados grupos de esponjas podem conter até 50% de sua biomassa de bactérias. 50%. Logo, é um animal ou uma colônia de bactérias?
Esta troca de genes entre espécies de diferentes grupos biológicos é chamada de transferência lateral de genes, ou transferência horizontal de genes, em oposição à transferência vertical de genes, que ocorre entre gerações de uma mesma espécie. Atualmente, se considera que a TLG é muito mais comum do que se pensava, e a árvore da vida de Darwin se transformou em uma rede de vida, na qual organismos vivos trocam genes de maneira promíscua.
Um exemplo recente foi a descoberta de que afídeos (insetos da ordem Hemiptera) apresentam capacidade de biossintetizar carotenos, e que os genes que codificam a biossíntese destes carotenos são muito parecidos com genes de fungos que também regulam a biossíntese de carotenos. Logo, tais organismos devem, em algum momento de sua história evolutiva, ter trocado genes que regulam a biossíntese de carotenos. Vamos à história.
Carotenos estão presentes em vários grupos biológicos da natureza: Archea (bactérias primitivas), bactérias, fungos e plantas. Por sua vez, os animais utilizam carotenos para diversas funções, como antioxidantes, como moduladores do sistema imune e como precursores para a formação de pigmentos da visão (em humanos, uma dieta muito pobre em carotenos causa uma doença chamada cegueira noturna). Mas, nenhum animal biossintetiza carotenos. Estes devem ser adquiridos através da dieta.
Os afídeos da espécie Acyrthosiphon pisum apresentam coloração amarela, alaranjada e até vermelha por conterem quantidades apreciáveis de carotenos. A coloração destes insetos está relacionada à sua predação (seus predadores têm preferência por afídeos de determinada coloração): afídeos verdes não são consumidos por predadores de afídeos vermelhos, e vice-versa. Os afídeos verdes apresentam alfa-caroteno, ß-caroteno e gama-caroteno, enquanto que os afídeos vermelhos apresentam os mesmos compostos e também toruleno e desidro-gama,psi-caroteno. A presença de tais carotenos em afídeos era creditada à sua dieta. Contudo, a composição carotenóide dos afídeos é significativamente diferente da composição carotenóide das plantas de que se alimentam, e tal hipótese passou a ser questionada. Uma segunda hipótese levantada é que os carotenos de afídeos teriam origem bacteriana, de seus endossimbiontes. Porém, a análise genômica dos endossimbiontes de A. pisum não indicou qualquer gene com homologia a genes que comumente codificam a biossíntese de carotenos (em plantas, por exemplo). Além disso, os endossimbiontes de A. pisum são transmitidos entre gerações pela fêmea, e, portanto, se determinada pelos endossimbiontes a distribuição de cores deveria seguir uma distribuição genética determinada pela mãe. Na verdade, as variações de cor entre os afídeos seguem uma distribuição mendeliana entre gerações, o que significa que os genes que codificam a biossíntese dos carotenos estão igualmente presentes nos machos e nas fêmeas dos afídeos.

Clones verdes (A), vermelhos (B) e clone verde mutante (C) obtido a partir do clone vermelho do afídeo A. pisum
Logo, os genes codificantes da biossíntese de carotenos em afídeos devem ser do genoma destes insetos.
Uma vez que o genoma de A. pisum já havia sido seqüenciado, a análise deste buscando genes codificadores de carotenos levou à descoberta de genes de carotenos muito similares de genes que codificam a biossíntese de carotenos em fungos. Assim, considerou-se a possibilidade da contaminação do DNA de A. pisum quando de seu seqüenciamento. Porém, a expressão de sete genes de A. pisum mostrou ser altamente reprodutiva em diferentes laboratórios de pesquisa. Logo, a contaminação também foi descartada.
Os dados obtidos indicam que houve transferência lateral de genes de um fungo para um afídeo em um evento único, seguido de duplicação do genoma do afídeo. Possivelmente, o doador do gene ancestral pode ter sido um fungo patogênico ou simbionte de afídeos, ou ainda um simbionte de plantas fonte de alimento de afídeos. A transferência lateral de genes também foi evidenciada para outra espécie de afídeo, Myzus persicae. O mais impressionante é que a variação de carotenos decorre de uma única substituição no gene, de guanosina para adenosina, que leva à substituição de um único aminoácido na enzima dessaturase carotenóide, de ácido glutâmico para lisina. Esta mudança altera totalmente o sítio ativo enzimático, já que o ácido glutâmico, como o próprio nome diz, é um aminoácido de caráter ácido, enquanto que a lisina é um aminoácido com caráter básico. Tal mudança no sítio enzimático determina a cor do caroteno produzido, e esta mutação está presente nesta enzima encontrada em bactérias, plantas e fungos. A mutação não permite mais a biossíntese de toruleno e desidro-gama,psi-caroteno, mas somente de gama-caroteno.
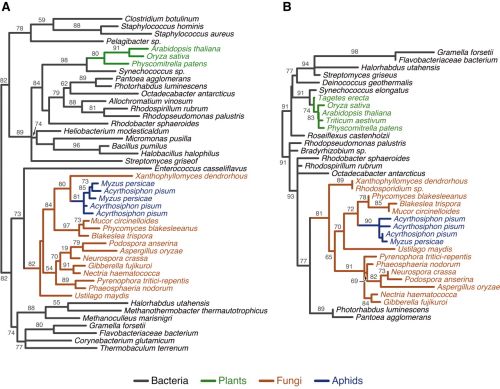
Árvores filogenéticas de enzimas que codificam a biossíntese de carotenos. (A) Dessaturases carotenóides e (B) cyclase carotenóide-sintase carotenóide. As sequencias ilustradas são de afídeos, bactérias, plantas e fungos. Não foram encontrados homólogos que codificam a biossíntese de carotenos em genomas de outros animais.
A história chega ao fim quando se verifica que a transferência lateral de genes codificadores da biossíntese de c
arotenóides de um fungo para um afídeo promoveu uma série de eventos, como duplicações genéticas, diversificação seqüencial e mudanças na expressão de cópias dos genes, que resultaram no surgimento de variantes de afídeos com diferentes cores. Suas diferentes cores promovem diferentes interações com o ambiente (absorção de luz) e com outros organismos (predadores). Levando-se em conta que carotenos são essenciais para muitas espécies de animais, é surpreendente que tal transferência lateral de genes não tenha ocorrido com mais freqüência. De qualquer forma, esta é a primeira aquisição de genes codificantes da biossíntese de carotenos por animais já documentada.
Moran, N., & Jarvik, T. (2010). Lateral Transfer of Genes from Fungi Underlies Carotenoid Production in Aphids Science, 328 (5978), 624-627 DOI: 10.1126/science.1187113
A conservação da biodiversidade é possível?
Segundo Escobar, na crescente perda de diversidade biológica “pesam os impactos cada vez mais severos das mudanças climáticas, da pesca predatória, da disseminação de espécies invasoras, do desmatamento e do consumo de recursos naturais (como água e solo) por uma população global cada vez maior e mais consumista.”
Aparentemente o Brasil tem atuado de maneira relativamente positiva, criando novas áreas de conservação e reduzindo a taxa de desmatamento. Segundo a reportagem, o Prof. Carlos Joly, do Instituto de Biologia da Universidade Estadual de Campinas, dá nota 7 para a atuação do Brasil junto à CDB.


foto: cortesia do Dr. Eduardo Hajdu (Museu Nacional, UFRJ)
O artigo de Escobar n’O Estado de São Paulo inclui uma análise de Matt Foster, Diretor da ONG Conservação Internacional e co-autor do trabalho publicado na Science. Segundo Foster, “Uma nova visão para a conservação da biodiversidade é necessária. Os líderes mundiais precisam ser mais ambiciosos nos seus esforços para proteger a natureza e todos os bens que ela nos provê via itens essenciais, como água limpa, ar puro, remédios e alimentos. Alguns países têm feito a lição de casa e estão adotando respostas adequadas, mas ainda há um longo caminho pela frente. O Brasil, por ser um dos países com a maior diversidade do mundo, tem a oportunidade e a responsabilidade de liderar o caminho.”
Este assunto já foi por mim abordado no blog “Química de Produtos Naturais” mais de uma vez (vejam aqui, aqui e aqui), e manifesto novamente aqui minhas observações. Na verdade, a questão da conservação da biodiversidade será realmente enfrentada somente quando o desenvolvimento econômico começar a ficar comprometido tanto para países desenvolvidos como para os países em desenvolvimento. Questões sobre conseqüências para a saúde, recuperação de áreas degradadas que têm impacto direto sobre as atividades humanas (como recifes de corais, por exemplo, que servem de berçário para várias espécies de peixes), sobre a importância em melhor se conhecer para se explorar de maneira racional os recursos naturais, são todas questões secundárias, que não estão incluídas na agenda da maioria dos governos e empresários. Mesmo daqueles que se dizem “defensores da preservação natural”.
A manutenção da biodiversidade, por si só, não interessa dirigentes políticos, empresários, banqueiros e outros setores da sociedade a favor do desenvolvimento a qualquer preço. A propaganda com “atitudes ecologicamente responsáveis”, “desenvolvimento sustentado”, “práticas verdes”, estão longe, muito longe de resultarem em ações efetivas que minimizem de fato a perda de biodiversidade e promovam não somente a manutenção como a recuperação de ecossistemas-chave para a manutenção da diversidade biológica. Tanto isso é verdade que os resultados estão disponíveis, no artigo de Escobar e no artigo publicado na Science.
Após 8 anos, o que se tem a dizer sobre a atual conservação da biodiversidade? Que é uma vergonha. Leia o artigo de Herton Escobar aqui.
Butchart, S., Walpole, M., Collen, B., van Strien, A., Scharlemann, J., Almond, R., Baillie, J., Bomhard, B., Brown, C., Bruno, J., Carpenter, K., Carr, G., Chanson, J., Chenery, A., Csirke, J., Davidson, N., Dentener, F., Foster, M., Galli, A., Galloway, J., Genovesi, P., Gregory, R., Hockings, M., Kapos, V., Lamarque, J., Leverington, F., Loh, J., McGeoch, M., McRae, L., Minasyan, A., Morcillo, M., Oldfield, T., Pauly, D., Quader, S., Revenga, C., Sauer, J., Skolnik, B., Spear, D., Stanwell-Smith, D., Stuart, S., Symes, A., Tierney, M., Tyrrell, T., Vie, J., & Watson, R. (2010). Global Biodiversity: Indicators of Recent Declines Science DOI: 10.1126/science.1187512
