


a culpa é dos cientistas? Em parte (II)
A segunda parte desta postagem se refere à divulgação científica propriamente dita.
A culpa é dos professores/pesquisadores que não se dedicam a esta atividade? Em parte.
Durante 3 anos (2009 – 2011) mantive um blog chamado “Química de Produtos Naturais”. Na época estava muito animado com a comunicação com estudantes e alguns poucos colegas através do blog. Lia muito, postava muito, respondia comentários, pesquisava, escrevia, entrei para o Science Blogs Brasil, além de fazer minha pesquisa, dar minhas aulas, escrever projetos, fazer análises de projetos, relatórios e artigos (revisão por pares), participar de comitês, organizar eventos, participar de ações de política científica, gerenciar os recursos do meu grupo (felizmente sem gerenciar a grana, diretamente, mas sim como gastar esta), a formação de meus alunos, interagir com os colegas, enfim, uma vida extremamente intensa.
Mas, aos poucos fui percebendo que minha atividade de divulgação científica no blog não tinha a menor importância. Muito pelo contrário. Aos poucos fui observando que não somente meus colegas liam, mas não comentavam (com poucas exceções), mas também os alunos. E mais: percebi que esta minha atividade estava virando motivo de chacota entre meus colegas acadêmicos. Um me disse “Não perco tempo lendo blogs. É totalmente inútil. Leio literatura científica.” Outro: “Faça isso, professor, que é isso o que você faz bem.” “OK”, pensei, “então paro por aqui”.

E parei.
Poucos colegas, muito poucos mesmo, se importaram com o fato de eu ter interrompido as atividades do blog “Química de Produtos Naturais”. A maioria nem percebeu (mesmo seguindo por RSS). Apenas alguns leitores assíduos perceberam.
Desde então, nunca deixei de acompanhar os blogs do Science Blogs Brasil, e outros como o Gene Reporter, o do Simon Schwartzman, o do Roger Pielke Jr. (excelente blog sobre política científica e outros assuntos, mas que também foi interrompido), o do The Breakthrough Institute, Polimerase de Mesa, Ciência na Mídia, o blog Scielo em Perspectiva, DNA Cético, Sobrevivendo na Ciência, Ciência Prática, Seed Magazine, Maurício Tuffani e Cultura Científica. Querem saber? Acho muito legal, e aprendo muito. E acho muito bacana mesmo que existam pessoas que trabalhem com divulgação científica de diferentes maneiras.
Já conversei com diversos colegas meus de outros países, dizendo que eu tive um blog. Ficaram muito (positivamente) surpresos, e pediram que eu escrevesse em inglês. Bom, não é tão fácil assim. Mas, fiquei espantado de ver como vários destes colegas estrangeiros acharam isso interessante.
Na crise atual em que se encontra a atividade científica no Brasil, todas estratégias são válidas para minimizar os estragos. Melhorar a gestão das universidades, institutos e centros de pesquisa. Tentar mesmo fazer pesquisa de melhor qualidade (o que não quer dizer simplesmente publicar em melhores revistas, de maior fator de impacto). Investir de verdade na formação dos alunos, levando muito a sério as atividades de docência. E divulgar o que está acontecendo nas universidades, em pesquisa, ensino e extensão.
Alguns sites de universidades são indecentes, de tão ruins. Às vezes é muito difícil encontrar o endereço de e-mail de um pesquisador/professor. As informações são pífias, tanto em forma quanto em conteúdo. Buscar informações sobre pesquisas, então, nem se fale. Eu diria que mais de 95% dos professores/pesquisadores não estão nem um pouco interessados em divulgar suas atividades para os próprios colegas e possíveis candidatos a alunos ou pesquisadores, quanto mais para a sociedade.
Entre 2013 e 2014 o programa BIOTA da FAPESP (também aqui) decidiu realizar uma série de palestras sobre biodiversidade, biomas do Brasil e outros aspectos ligados à pesquisa e conhecimento desta natureza, uma iniciativa chamada BIOTA Educação (veja que legal, aqui). Foi um sucesso absoluto. O auditório da FAPESP ficou muitas vezes lotado de gente que ia assistir as palestras, muitas vezes alunos de escolas.

Estou absolutamente convencido que a sociedade QUER saber mais sobre pesquisa. QUER mais conhecimento e mais informação científica. Mas, não basta os professores/pesquisadores se engajarem nestas atividades se estas não forem minimamente reconhecidas pelos pares. As próprias instituições tem que reconhecer tais esforços e estimular aqueles que se engajam nestas atividades. Seria MUITO LEGAL que fosse possível obter algum financiamento, mesmo pequeno, para se escrever um blog, escrever textos e livros de divulgação científica, participar de atividades culturais, fóruns de discussão sobre este assunto, etc.
Os pesquisadores, as instituições de ensino e pesquisa, os órgãos de fomento, os governos, todos falam muito do problema da educação no país, que é muito sério. Penso que, se houvesse mais divulgação científica, e esta fosse valorizada e reconhecida, a sociedade em geral iria querer se educar cada vez melhor, para entender cada vez melhor o mundo à sua volta.
Milagres não existem. Por melhores que sejam os governos, administradores, professores/pesquisadores, se não houver um trabalho conjunto para se melhorar o nível da educação e do conhecimento científico da sociedade em geral, esta mudança será muito mais lenta.
A culpa é dos cientistas? Em parte. (I)
O Átila me provocou. De verdade. Tanto que voltei, primeiro para comentar a postagem dele. Depois para… bem, escrever de vez em quando. Com alguma frequência.
Voltando ao Átila, a postagem “A ciência brasileira está em crise e a culpa é dos cientistas” no Rainha Vermelha é muito provocativa. Por vários motivos, concordo com ele em alguns pontos e discordo em outros.
Primeiro ponto: universidades sem financiamento. As universidades públicas estão com financiamento reduzido. O orçamento da USP em 2016, por exemplo, será de cerca de R$ 4,6 bilhões. Dividindo por 12, isso dá cerca de R$ 383 milhões por mês. Esta grana está sendo destinada quase que exclusivamente para os salários. Como é o caso da UNICAMP e da UNESP. Como é o caso de todas as outras universidades públicas do Brasil. Apenas em parte a culpa é dos professores/pesquisadores.
Isso porque alguns destes tentam ser heróis, com a melhor das boas intenções. Administrar os institutos, faculdades, centros, museus, etc., não é brincadeira, não. É coisa para profissional. Na minha opinião já passou da hora de se rever o modelo de administração das universidades públicas, pelo bem destas. E, na minha opinião, os professores/pesquisadores das universidades públicas deveriam entender que é muito difícil gerenciar, além de fazer pesquisa e dar aulas, e tentar fazer entender os governos que as universidades deveriam ser em parte administradas por profissionais. É minha opinião.

Vou citar um único exemplo.
Entre julho de 2014 e março deste ano fiz um estágio sabático no Life Sciences Institute da Universidade de Michigan, em Ann Arbor, EUA. Enquanto boa parte das universidades públicas estaduais dos EUA passa por sérias dificuldades financeiras, não é este o caso da Universidade de Michigan (UM). A UM é uma universidade estadual, financiada em parte pelo governo do estado de Michigan. Classificada em posição 17 no último ranking Times Higher Education – o mais prestigioso ranking de universidades – a UM atingiu status de uma das melhores universidades do mundo ao longo dos primeiros 15 anos do século 21. As razões para isso são várias.

No início dos anos 2000 a administração da universidade solicitou a um grupo de “experts” uma análise das variáveis financeiras que poderiam afetar a economia da universidade nos anos seguintes. Acertadamente o grupo previu que a indústria automobilística – a maior fonte de renda do estado de Michigan até 2008 – não iria ter para sempre a mesma pujança. A partir de então a Universidade de Michigan buscou diversificar a busca de recursos. Aumentou os valores das anuidades estudantis, por um lado. Por outro, buscou doadores. E tratou de implementar uma gestão eficiente de seus recursos, atualmente considerada modelo no sistema universitário norte-americano.
Desde 1970 a dependência do orçamento da UM do governo estadual diminuiu de 64% para 16%. A importância das anuidades para o orçamento da universidade passou de 26% para 71%. Ao longo da última década, o dispêndio anual por aluno do governo do estado de Michigan caiu de US$ 10.579,00 para 6.753,00. A busca de doadores para a UM possibilitou duplicar o fundo de reserva da universidade de US$ 4,4 bilhões para quase US$ 9 bilhões. Apenas no último triênio (2012-2014) a UM arrecadou mais de US$ 2 bilhões em doações. Com tal fundo de reserva, a UM se posiciona apenas após as universidades de Harvard, Yale, Stanford e Princeton. Atualmente o fundo de reserva para todos os campi da Universidade da Califórnia não passa de US$ 7 bilhões. A diferença é enorme: todos os campi da Universidade da Califórnia incluem quase 400.000 estudantes. A Universidade de Michigan não tem 41.000 alunos.
Diferentemente das universidades brasileiras, não existem eleições para nenhum dos cargos da UM: reitor, pró-reitores, diretores de faculdades, cargos administrativos ou chefes de departamento são todos indicados e contratados, com base em experiência e competência. Assim, as atividades acadêmicas se concentram quase que exclusivamente no ensino, na pesquisa e nas atividades culturais.
Além de 22 prêmios Nobel dentre ex-estudantes, ex-professores e professores, a UM é um verdadeiro patrimônio da cidade de Ann Arbor, de pouco mais de 110.000 habitantes. Crescendo com a universidade ao longo de praticamente 200 anos, a cidade e a universidade são quase uma só. Ann Arbor tem população com alto nível educacional. É possível não fazer nada além de visitar museus, assistir palestras, seminários, participar de eventos culturais os mais diversos. Os principais teatros da cidade são da universidade, como o Hill Auditorium que na temporada 2014-2015 teve artistas como Itzhak Perlman, Sinfônica de San Francisco, Wynton Marsalis, Academy of St. Martin in the Fields e Gilberto Gil (veja aqui a programação da temporada 2015-2016).

O estádio da UM, conhecido como “The Big House”, é o maior estádio aberto dos EUA, com capacidade para quase 110.00 pessoas, o segundo maior do mundo. Se lotado, a cada jogo se arrecada quase US$ 10 milhões somente em ingressos, sem contar os direitos de transmissão por TV. Manchester United x Real Madrid jogaram no estádio da UM em setembro de 2014, com mais de 100.000 ingressos vendidos em menos de 24 h.

O sistema de transporte público de Ann Arbor é considerado modelo. Sem surpresas, a UM financia 40% do mesmo, pagando integralmente as passagens de seus estudantes e funcionários (inclusive visitantes :-)). Com uma qualidade de vida excelente, polo cultural e de inúmeras companhias de inovação e desenvolvimento tecnológico, Ann Arbor é centro de atração de jovens talentos. Ao final de 2014, duas vagas para professor oferecidas pelo Life Sciences Institute da UM tiveram 200 candidatos com os melhores CVs de jovens pesquisadores dos EUA.

Lojas e mais lojas vendem produtos com o logo UM. Orgulhosamente a população de Ann Arbor os ostenta. Não sem razão. A Universidade de Michigan é muito mais do que uma universidade. É uma marca.

Razão? Excelentes ensino e pesquisa, resultado de excelente gestão.
Continua…
Fatores reguladores da virulência de Staphylococcus aureus
Antibióticos, bactérias, infecções, microrganismos são alguns dos assuntos que mais gosto, como meus leitores já devem ter percebido. Já foram postados vários textos neste blog sobre esta temática. Também já escrevi aqui sobre a bactéria patogênica Staphylococcus aureus, que pode ser extremamente nefasta, principalmente se a infecção por esta não for adequadamente tratada.S. aureus, como é conhecida, é uma bactéria danada. Ela possui vários mecanismos de virulência, como proteínas que atuam na adesão desta bactéria a superfícies e na invasão de tecidos de hospedeiros, exoproteínas que atuam contra mecanismos de imunidade, além de também possuir toxinas que causam hemólise (destruição dos hemócitos, ou glóbulos vermelhos) e que formam poros em membranas, permitindo o “vazamento” de íons e outras moléculas pequenas de células de hospedeiros. Para que cause infecção, é necessário que os fatores de virulência de S. aureus atuem de forma coordenada, até mesmo de maneira redundante, de tal forma que se um de seus fatores de virulência for afetado, os outros continuarão ativos.
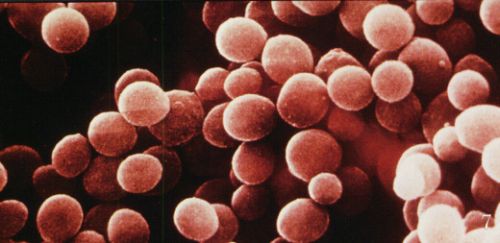
Staphylococcus aureus
Descobriu-se que S. aureus tem um gene regulador de virulência, denominado agr (accessory gene regulator). Até pouco tempo atrás se sabia que a expressão do gene agr era regulada por um sinalizador químico. E se sabia de que tipo era este sinalizador: uma molécula pequena, da classe dos metabolitos secundários. Também se descobriu que, apesar do gene agr ter uma grande importância na regulação da virulência de S. aureus, este gene não é essencial para a expressão da virulência. São conhecidos isolados clínicos (de hospitais) de S. aureus desprovidos do gene agr.Recentemente, pesquisadores realizaram uma extensa análise do material genético (genoma) de S. aureus, de maneira a verificar a presença de genes reguladores da biossíntese (processo enzimático de várias etapas que é responsável pela formação das moléculas biológicas, como proteínas, açúcares, DNA, RNA, e também de metabolitos secundários) de peptídeos não-ribossômicos exclusivos de S. aureus. Analisando o genoma desta maneira, é possível se detectar a presença de genes que regulam a biossíntese de substâncias específicas, e propor a estrutura química das mesmas, uma vez que estes genes coordenam a ação de enzimas que atuam de maneira bem estabelecida. Foram analisados 50 genomas de S. aureus, e identificado um agregado de genes (gene cluster) responsável pela biossíntese de peptídeos não-ribossomais (ou seja, peptídeos que não são formados nos ribossomos, como são a grande maioria das proteínas, peptídeos muito grandes) ainda desconhecidos. O agregado de genes mostrou ter um gene de 7,17 Kb (1 Kb = 1000 bases nucléicas, que formam o DNA), que ocupa apenas 0,25% do genoma de S. aureus.
Este agregado de genes regula a formação de uma sintetase de peptídeos não-ribossomais (NRPS, non-ribossomal peptide synthetase) de 2389 aminoácidos. Esta NRPS mostrou ter sítios que atuam na formação de um dipeptídeo (um derivado de dois aminoácidos) com estrutura muito peculiar: é formado a partir dos aminoácidos valina e tirosina, que se condensam e dão origem a uma estrutura cíclica, de massa molecular de 262,17 u.m.a.(u.m.a. = unidade de massa atômica). Ou seja, a partir da análise do genoma de S. aureus, foi possível se encontrar genes que regulam a formação deste dipeptídeo, cuja estrutura pôde ser predita a partir dos resultados obtidos da análise do genoma.
Porém, era necessário que esta predição fosse confirmada.
Desta forma, os autores deste estudo cresceram S. aureus em meio de cultura, extraíram o meio de cultura com solventes orgânicos e analisaram os extratos do meio de cultura por cromatografia líquida (HPLC, high-performance liquid chromatography) acoplada a um detector de espectrometria de massas. Desta forma, para cada substância separada foi possível obter um espectro de massas, que indica a massa molecular da forma protonada das substâncias presentes no meio de cultura de S. aureus. Estas análises indicaram a presença de duas substâncias (dentre outras) no extrato orgânico do meio de cultura de S. aureus: uma de massa 262 e outra de massa 246. As duas mostraram ser muito parecidas pelo fato de serem pouco separadas na análise por HPLC, e também por apresentarem espectro no ultravioleta muito similar. Ou seja, talvez estas substâncias apresentassem estruturas muito parecidas, e tivessem uma biossíntese comum.
Esta hipótese foi confirmada após a obtenção das substâncias puras, uma separada da outra. Após isoladas, puderam ser analisadas por várias técnicas espectroscópicas, principalmente por diferentes experimentos de ressonância magnética nuclear (RMN). Estas análises indicaram que um dos dipeptídeos isolados era formado por tirosina e valina (aureusimina A), e o outro por fenilalanina e valina (aureusimina B). A diferença entre os dois é de apenas um átomo de oxigênio.
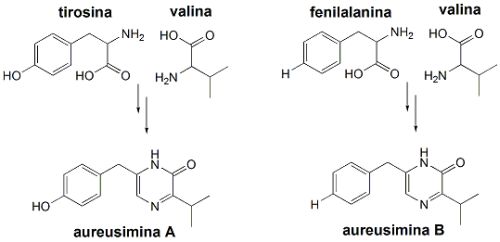
Para confirmar que estes dois dipeptídeos eram realmente biossintetizados por S. aureus, o gene ausA, supostamente responsável pela regulação da biossíntese destas duas substâncias, foi substituído por um gene artificial. E S. aureus não mais produziu as aureusiminas.Em seguida, foram feitos testes com as duas substâncias (aureusiminas A e B) para verificar como estas influenciavam na expressão do gene agr, que controla a virulência de S. aureus. Observou-se que a substituição do gene ausA de S. aureus, responsável pela biossíntese das auresiminas, por um gene artificial alterou completamente o padrão de expressão do gene agr, responsável pela virulência. Observou-se diminuição na formação de proteínas imunomodulatórias, de proteínas de adesão, bem como de proteínas líticas (que promovem o rompimento de membranas) e de citotoxinas (toxinas de células). Enquanto tais proteínas mostraram ter sua produção estimulada na presença das aureusiminas, na ausência destas substâncias as proteínas virulentas mostraram ser produzidas em quantidades muito menores.
Claramente as aureusiminas mostraram atuar na regulação das proteínas participantes do processo de virulência de S. aureus.
Normalmente, S. aureus “original”, sem ter seu genoma alterado, promove hemólise. Quando o gene ausA foi suprimido, a hemólise promovida por S. aureus sem este gene também foi suprimida. Quando se adicinou as aureusiminas no meio de crescimento de S. aureus com seu genoma alterado (com o gene ausA deletado), voltou-se a observar a hemólise. Claramente as aureusiminas participam diretamente da ativação dos processos de virulência de S. aureus.
E mais: quando camundongos sadios foram infectados com S. aureus “original” (sem ter seu genoma alterado), os bichinhos apresentaram grande quantidade de unidades formadoras de colônia (um indicativo de proliferação bacteriana) em seus rins, fígado, baço e coração. Quando os camundongos foram infectados com S. aureus mutada, com o gene ausA suprimido, observou-se que os rins apresentaram grau de infecção, mas os outros três órgãos muito menos, principalmente o coração (praticamente sem infecção). Ou seja, a inibição da biossíntese das aureusiminas levou a um grau de infecção virulenta muito menor.
A descoberta que as aureusiminas controlam a virulência de S. aureus abre um enorme conjunto de possibilidades para o desenvolvimento de novas formas de tratamento de infecções por esta bactéria. Por exemplo, a eventual descoberta de inibidores da biossíntese das aureusiminas pode significar a abolição do uso de antibióticos para o tratamento de infecções por S. aureus. Embora tal perspectiva possa parecer distante, tudo depende do esforço de pesquisadores acadêmicos e de indústrias farmacêuticas sérias. O mecanismo de ação de tais agentes inibidores da virulência de S. aureus seria completamente diferente do mecanismo de ação dos antibióticos. Isso porque os inibidores da biossíntese das aureusiminas interferem no processo bioquímico de formação destas substâncias, muito mais difícil de sofrer uma mutação benéfica para continuar promovendo a virulência. Desta forma, não seria mais necessário utilizar antibióticos, dos quais as bactérias adquirem resistência (através de mutações) em poucas gerações.
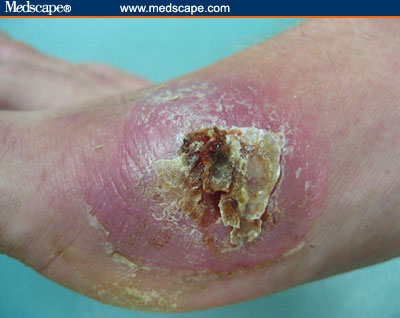
Infecção causada por S. aureus (nojenta!)
Embora o trabalho realizado com S. aureus pareça muito complicado, realmente não é. As atuais ferramentas bioquímicas permitem a realização de um trabalho deste em tempo relativamente curto. Na verdade, os autores mencionam um trabalho publicado em 2008 como fonte das informações preliminares nas quais se basearam para realizar seu estudo. A referência original indica que este trabalho (Novick e Geisinger) foi publicado on-line em agosto de 2008. Ou seja, o estudo realizado com S. aureus levou, no máximo, menos de dois anos para ser desenvolvido. Uma boa idéia e abordagens e estratégias de trabalho bem delineadas levaram à publicação de um artigo extramemente relevante na Science, uma vez que as infecções causadas por S. aureus são consideradas um dos maiores problemas de saúde pública no mundo.O que certamente fez a diferença para um trabalho deste ter sido feito em tão pouco tempo? Acesso a instrumentação e aos materiais (reagentes, linhagens bacterianas, camundongos) necessários para o desenvolvimento deste projeto. Sem entraves, sem burocracia, sem demora, sem falta de dinheiro.
 Wyatt, M., Wang, W., Roux, C., Beasley, F., Heinrichs, D., Dunman, P., & Magarvey, N. (2010). Staphylococcus aureus Nonribosomal Peptide Secondary Metabolites Regulate Virulence Science, 329 (5989), 294-296 DOI: 10.1126/science.1188888
Wyatt, M., Wang, W., Roux, C., Beasley, F., Heinrichs, D., Dunman, P., & Magarvey, N. (2010). Staphylococcus aureus Nonribosomal Peptide Secondary Metabolites Regulate Virulence Science, 329 (5989), 294-296 DOI: 10.1126/science.1188888
Novick, R., & Geisinger, E. (2008). Quorum Sensing in Staphylococci Annual Review of Genetics, 42 (1), 541-564 DOI: 10.1146/annurev.genet.42.110807.091640
A química supramolecular das aranhas
O fio das teias de aranhas é um material absolutamente excepcional: tem uma resistência mecânica proporcionalmente maior do que a do aço quando se leva em conta sua baixa densidade. Um único fio de teia de aranha alinhado sobre a linha do equador teria a massa de apenas 500 gramas. Os fios da teia de aranhas são feitos de proteínas: uma cadeia polipeptídica que consiste de uma sequência repetida de dois fragmentos peptídicos, juntos chamados de “sequência AQ”. O fragmento A é hidrofóbico (hidro=água; fobia=aversão. Ou seja, aversão à água, um fragmento que não admite moléculas de água associadas), que apresenta muitas unidades do aminoácido alanina (A): GPYGPGASA6GGYGPGGQQ (cada letra corresponde a um aminoácido). O fragmento Q é hidrofílico (hidro=água; filo=afinidade. Ou seja, com afinidade por água, é um fragmento que apresenta moléculas de água associadas através de ligações de pontes de hidrogênio), rico em glutamina e em glicina: (GPGQQ)4. Esta estrutura AQ é a “parte central” da fibra do fio da teia de aranha, e sua estrutura e função lembram a do colágeno (proteína espalhada pelo corpo dos humanos que forma o tecido conjuntivo). A fibra dos fios da teia de aranha são formados por 12 fragmentos AQ, os quais terminam com fragmentos diferentes, que atuam como “sinais químicos” e partes diferentes constituídas por grupos amina livres (-NH2) e grupos carboxila lvres (-CO2H). Estas partes diferentes formadas pelos grupos amina e carboxila livres participam de funções diferentes, como controle da solubilidade da proteína e da formação da fibra que dá origem ao fio da teia de aranha.
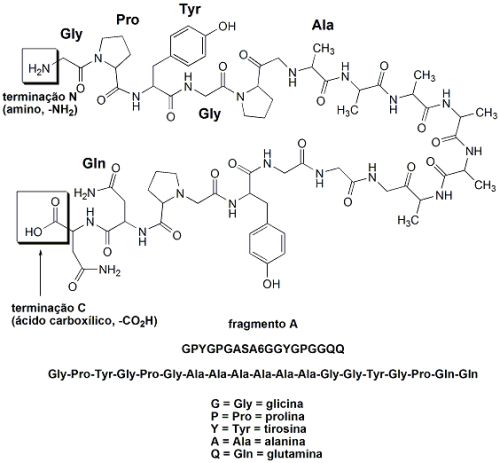
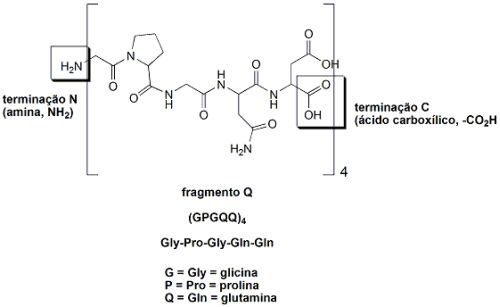
Por exemplo, a estrutura da terminação C (ácido carboxílico, -CO2H) não repetida (NR-C) da fibroína da aranha Aaraneus diadematus foi estudada em solução por ressonância magnética nuclear (RMN). A técnica de RMN detecta os núcleos de alguns átomos (mas não de todos), como por exemplo os de hidrogênio (1H). É possível se “medir” a forma como átomos de hidrogênio de uma molécula orgânica interagem entre si, e com isso entender como esta molécula orgânica se comporta em solução (por exemplo, em uma solução salina). Desta forma, é possível se verificar como partes das proteínas das aranhas “se dobram” em solução (foi o que foi feito). Foi observado que dois NR-C da fibroína de A. diadematus formam uma estrutura dimérica, altamente simétrica, com um formato de barril, constituído por duas hélices da proteína, unidas entre si por uma ponte dissulfeto (pontes dissulfeto, S-S, são ligações enxofre-enxofre formadas entre dois aminoácidos cisteína. Estas pontes dissulfeto aumentam a rigidez das proteínas). Este “formato de barril” da estrutura das proteínas das fibras dos fios da teia fazem com que as partes hidrofóbicas (A) fiquem agregadas, enquanto que a parte hidrofílica (Q) fica fora do barril, de maneira a que possa se agregar a moléculas de água.
Para que o fio da teia seja formado, é necessário que o pH do ambiente de formação seja muito bem regulado. Foram feitos experimentos com mini-spindroínas (proteínas bem menores do que as spindroínas, que formam os fios), que mostraram que quando a terminação N (amino, -NH2) está participando da formação da proteína, o pH do ambiente deve ser 6,0 (levemente ácido).De outra forma, em pHs mais baixos (mais ácidos) ou maiores (mais básicos), a proteína não se forma. Todavia, estes efeitos são reversíveis. Ou seja, se por um acaso o pH do ambiente se altera, e a formação da proteína é interrompida, a formação da proteína continua quando o pH do ambiente volta a ser 6,0. Já a terminação C (ácido carboxílico, -CO2H) participa da extensão da proteína, ou seja, do aumento do tamanho desta.
A formação das proteínas do fio da teia ocorre dentro de glândulas das aranhas, e são estocadas como oligômeros (pequenos polímeros) de alta densidade, com uma estrutura micelar (uma micela é uma estrutura esférica supra-molecular, ou seja, que faz uso de muitas moléculas de um determinado tipo, chamadas de detergentes, por terem uma extremidade carregada positiva ou negativamente, hidrofílica, e uma “cauda” sem carga, hidrofóbica), formando uma microemulsão. Cada micela contém várias proteínas dos fios da teia, e as micelas são estabilizadas pela presença dos grupos N (amina, -NH2) e C (ácido carboxílico, -CO2H) terminais. A auto-agregação das proteínas para formar as micelas é reversível.
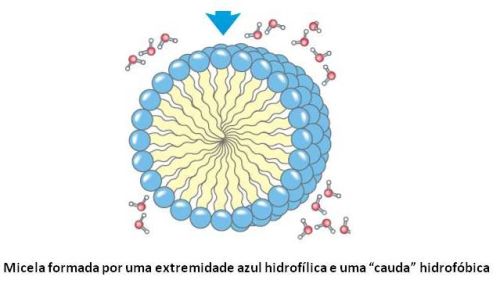
Dentro das aranhas, a formação das fibras de proteínas não é apenas determinada pelas mudanças químicas do ambiente, mas também por estímulos mecânicos em um tipo de sinergia (ação conjunta de forças) de efeitos macroscópicos e microscópicos, ilustrados na figura a seguir. Para entender as propriedades formadoras de fibrilas da cadeia de proteínas, é preciso compreender como a extrusão do complexo micelar supra-molecular acontece: estresse de cisalhamento (cisalhamento é um movimento de “escorregar para frente e para trás” ao longo do comprimento de um eixo), extrusão de água e troca de íons estimulam a formação das fibras dos fios da teia. A terminação C (ácido carboxílico) faz com que a cadeia AQ não mude de viscosidade durante o processo de cisalhamento. Por outro lado, a cadeia protéica já formada dá origem a uma forma globular que leva ao surgimento de agregados fibrosos quando o terminal C (ácido carboxílico) está presente.
As aranhas formam uma proteína extremamente específica para dar origem às fibras dos fios da teia. Estas proteínas são o resultado da fusão de um bloco de um co-polímero (AQ) anfifílico (anfi=ambos; filo=afinidade. Ou seja, um polímero que é ao mesmo tempo hidrofílico e hidrofóbico) e dois “barris” diméricos, com proteínas dobradas na forma de alfa-hélices. Nas glândulas da aranha, as proteínas formam um conjunto de uma microemulsão supramolecular. A aranha então aplica fortes forças de cisalhamento ao mesmo tempo que realiza a extrusão da proteína de suas glândulas. Esta extrusão é acompanhada da expulsão de água da microemulsão, levando ao desdobramento das estruturas de barris, fazendo com que os fragmentos hidrofílicos se projetem de dentro para fora. Ocorre então uma mudança macroscópica na forma da emulsão, levando a uma polimerização específica que dá origem aos fios da teia com sua enorme resistência mecânica.

A transição na forma das proteínas dos fios da teia, a partir de um complexo supramolecular solúvel em água na forma de micelas para formar fibras longas, mostra como estas proteínas podem mudar de forma de acordo com mudanças no ambiente químico e sob forças mecânicas. Além disso, o estudo realizado mostrou não somente as particularidades de como os fios das teias de aranha e como a teia é formada, mas também como as aranhas trabalham com os fios para formar a teia mais eficiente para capturar suas presas. Embora a formação de fibrilas protéicas sempre foi vista co
mo uma disfunção estrutural, levando à perda de forma tridimensional das proteínas que as formam, as aranhas se utilizam de tais propriedades para formar os fios de suas teias, altamente resistentes.
Perguntas que ainda restam para serem respondidas sobre as teias de aranha incluem: será que as terminações N (amina) e C (ácido carboxílico) operam de maneira cooperativa, ou reagem diferentemente sob um determinado estímulo químico? Como a agregação supramolecular das cadeias protéicas individuais garante a formação de uma estrutura em rede para dar origem a fios de até 1 metro de comprimento? Como que um fio formado de maneira tão específica pode responder tão bem a diferentes presas capturadas pelas aranhas? A qualidade dos fios das teias depende de como esta é formada pela aranha e fatores ambientais, bem como da alimentação das aranhas. Mas a qualidade intrínseca dos fios é determinada pela sua composição de aminoácidos. A compreensão de como os componentes dos fios da teia pode dar origem ao processo de formação da teia, com suas propriedades mecânicas, ainda permanece um enigma para as ciências dos biomateriais.
Só o homem-aranha conhece estes segredos.
Silvers, R., Buhr, F., & Schwalbe, H. (2010). The Molecular Mechanism of Spider-Silk Formation Angewandte Chemie International Edition DOI: 10.1002/anie.201003033
Quanta vida na Terra?
Mas May também gosta de apreciar as brincadeiras alheias com números deste tipo. E de fornecer dados sem citar referências. Em seu mais recente artigo, “Tropical Arthropod Species, More or Less?”, May inicia suas divagações sobre quais impressões teriam extra-terrestres ao chegar na Terra. Segundo ele (May), estes teriam como primeira curiosidade saber quantas formas distintas de vida existem na Terra. Embora eu não concorde com este questionamento de May (acho que extra-terrestres teriam mais curiosidade em saber que TIPOS de vida existem na Terra), entendo a introdução de seu artigo como uma provocação sobre a nossa (humanos, terráqueos) própria ignorância sobre este assunto. Afinal, ao afirmar que “um terço dos taxonomistas trabalham com vertebrados (1% de todas as espécies biológicas), outro terço dos taxonomistas com plantas (cerca de 10% de todas as espécies biológicas), e o último terço com invertebrados (cerca de 90% de todas as espécies)”, May está querendo dizer que há pouca gente para conhecer tanta diversidade biológica. May acredita que o número total de espécies biológicas classificadas seja de 1,6 a 1,7 milhões, com cerca de 15.000 a mais a cada ano. Ao se levar em conta um número (bem conservador) de 3 milhões de espécies vivas, no total, estamos longe de conhecer a diversidade biológica da Terra.
May se detém na análise realizada por Hamilton e seus colaboradores, que se utilizou de estimativas e abordagens estatísticas modernas para estimar o número de artrópodes da Terra. Segundo a Wikipédia,
Os Artrópodes (do grego arthros: articulado e podos: pés, patas, apêndices) são animais invertebrados caracterizados por possuírem membros rígidos e articulados. São o maior grupo de animais existentes, representados pelos gafanhotos (insetos), aranhas (arachnida), caranguejos (crustáceos), centopeias (quilópodes) e embuás (diplópodes), somam mais de um milhão de espécies descritas (apenas mais de 890.000 segundo outros autores). Mais de 4/5 das espécies existentes são Artrópodes que vão desde as formas microscópicas de plâncton com menos de 1/4 de milímetro, até crustáceos com mais de 3 metros de espessura.

Hamilton e seu time partiram das idéias de Erwin, que tentou estimar o número de espécies de besouros (o maior grupo dos artrópodes) depois de realizar um estudo da distribuição vertical destes animais sobre uma única espécie de árvore tropical. Erwin estimou o número de espécies de artrópodes tropicais entre 30 e 100 milhões. A escolha de Erwin baseou-se no fato que a diversidade de artrópodes, em particular de besouros, nas regiões tropicais é muito maior do que em outras regiões da Terra. Já o time de Hamilton realizou vários estudos sobre a distribuição de besouros e outros artrópodes em árvores de diferentes países, como a Nova Guiné (na África; ou seria Papua Nova Guiné, no Pacífico Sul, perto da Indonésia?), Brasil, Panamá, Venezuela, Sulawesi, entre outros países.
Erwin estimou que 40% dos artrópodes seriam besouros. Os estudos do time de Hamilton levaram à conclusão, utilizando técnicas modernas e sofisticadas de análise estatística, que o número de besouros corresponderia de 22 a 60% dos artrópodes. Na média, muito próximo do resultado de Erwin. Este estimou que o número aproximado de árvores tropicais seria de 50.000. Hamilton e seus colaboradores chegaram a uma estimativa entre 43.000 e 50.000. A principal diferença nos resultados dos dois estudos, publicados com um intervalo de 28 anos, é que os besouros parecem ser muito menos especializados (segundo Hamilton) do que pareciam ser (segundo Erwin). Ou seja, besouros podem se alimentar em várias árvores de espécies diferentes, e não sempre na mesma e única espécie de árvore. Desta forma, o número de espécies de artrópodes estimado pelo grupo de Hamilton é bem menor do que entre 30 e 100 milhões (estimado por Erwin). Segundo Hamilton, este número estaria próximo de 3,7 milhões de espécies, com 90% de probabilidade de estar entre 2,0 e 74 milhões de espécies, quando se utilizou de um primeiro modelo de análise. Para um segundo modelo de análise, o número foi estimado de 2,5 milhões de espécies de artrópodes, com 90% de probabilidade de se situar entre 1,1 e 5,4 milhões de espécies (de artrópodes).
De acordo com os resultados apresentados por Hamilton (e discutidos por May), cerca de 2/3 das espécies de artrópodes da Terra ainda não foram descritas. Segundo May, este número seria um reflexo da distribuição desigual de taxonomistas para a descrição dos diferentes grupos de organismos vivos. Mas por outro lado, também que as agências financiadoras consideram taxonomia uma “ciência menor, apenas descritiva, e pouco geradora de conhecimento associado”. Por isso que a taxonomia é uma ciência em vias de extinção. E, em paralelo, as espécies biológicas também, mas não sabemos ao certo quantas nem quais.
May, R. (2010). Tropical Arthropod Species, More or Less? Science, 329 (5987), 41-42 DOI: 10.1126/science.1191058
Cientistas e mídia
Segundo a reportagem, “O pesquisador da Universidade da Louisiana Edward Overton já publicou uma pesquisa com o título ‘Eficiência da Fitorremediação e da Biorremediação de n-Alcanos como Função do Comprimento da Cadeira de Carbono em Ambientes Banhados’. Ele também é detentor da patente de algo chamado cromatografia de microestrutura com coluna retangular. E, recentemente, o professor emérito apareceu num talk-show campeão de audiência para explicar o petróleo aos leigos. Overton é um de dezenas de cientistas que trabalharam por anos longe da atenção do grande público e que agora se veem nos holofotes da mídia, tentando explicar o vazamento de óleo no Golfo do México. ‘Geralmente passo meu tempo analisando amostras e olhando para gráficos, o que não é muito sexy’, disse Overton, rindo. ‘Quem teria pensado que (David) Letterman iria convidar a mim, um cientista, para seu programa?’ Overton crê que é chamado pela mídia porque põe a questão “em linguagem clara”. Durante sua participação no show de Letterman, levou uma garrafa de óleo recolhido do golfo. Mas a fama vem com um preço. Ele passou a receber telefonemas de um matemático amador que acredita ter a solução do vazamento, e que rastreou Overton depois de vê-lo na televisão.”
Reparem no texto do artigo d’O Estado. Que informação útil se agrega a este se divulgando o título (aparentemente muito complicado) de trabalho anterior realizado pelo cientista? E ainda, no tom irônico quando se diz que “o pesquisador é detentor da patente de algo chamado cromatografia de microestrutura com coluna retangular”.
Continuando, ainda segundo a reportagem, Susan Ustin, professora de ciências do meio ambiente da Universidade ad Califórnia, Davis, teria declarado: “A maioria de nós não somos as pessoas mais extrovertidas do mundo. Não estamos acostumados a ser o centro das atenções, e a maioria de nós teme se enrolar”. Ela acrescenta que, no meio acadêmico, bater papo ao vivo na televisão e promover o próprio trabalho costuma ser visto com maus olhos. “É uma espada de dois gumes. Fico preocupada que vou dizer algo de uma certa maneira que, quando entrar na reportagem, não vai soar inteligente”, disse ela. “E fico preocupada que meus colegas não venham a aprovar”.
Intrigado com esta reportagem, comecei a pensar se cientistas não gostam da mídia porque via de regra a mídia deturpa informações científicas, porque esta só gosta de assuntos “quentes”, ou faz com que os cientistas não se sintam bem frente aos colegas. Ou por mais algum motivo? Ou seriam todos?

O fato é que a mídia colabora pouco com a ciência, no sentido de divulgar esta de forma correta, sem tentar misturar alhos com bugalhos. Por exemplo, recentemente Rafael Soares, biólogo formado pela UNESP – Rio Claro e doutorando pela Biotecnologia da USP na área de terapia gênica do câncer, foi convidado para participar do programa SuperPop de Luciana Gimenez para discutir do propalado “fim do mundo em 2012”. De cara a apresentadora fez a seguinte pergunta para Rafael: “Você é cético? Então não acredita em Deus?” Danou-se tudo. Qual é a relação que existe entre ser cientista e cético, ou entre ser cientista e não acreditar em Deus, ou entre ser cético e não acreditar em Deus? Nenhuma. Mas, de início Luciana Gimenez apenas reforçou um estereótipo comum, de que cientistas são céticos e não acreditam em Deus. Segundo Rafael, no decorrer do programa, quase nada se discutiu sobre o caráter realmente científico do “fim do mundo de 2012”.
Aparentemente existe uma enorme dificuldade na comunicação entre cientistas e a mídia. Êrros sobre resultados científicos divulgados são extremamente comuns (veja por exemplo, aqui). A mídia tem uma pressa exagerada em publicar um assunto porque acha que é “quente”, e acaba escrevendo textos pouco informativos e com bom grau de sensacionalismo, o que não ajuda. Até mesmo na televisão. A impressão que se tem é que o interesse em divulgar ciência aumentou (e muito), mas sem o devido cuidado por parte dos meios de comunicação.
Por outro lado, muitos cientistas acham que a mídia deturpa as informações, e por isso não gostam e não querem dar entrevistas. Neste caso, a má vontade só piora o que já está ruim. É necessário que haja uma aproximação dos cientistas junto aos veículos de informação. Isso porque a ciência é a chave para entender muito sobre o mundo à nossa volta. E o telespectador, o leitor leigo, busca informações e conhecimento.
Um texto de 2006 do jornal The Times/The Sunday Times cita o ex-primeiro ministro inglês Tony Blair, segundo o qual a Inglaterra precisava de cientistas midiáticos para inspirar a juventude. O texto fala da importância de assuntos como pesquisas com células-tronco, a utilização de energia nuclear, experimentos com animais, a utilização de sementes geneticamente modificadas para aumentar a produção de alimentos, mudanças climáticas, e que os jovens deveriam seguir carreiras científicas para contribuir para o bem social. Mas, será que ser um “cientista midiático” para divulgar ciência e chamar a atenção dos jovens é tão importante assim?

Talvez, de certa forma, sim. Mas, quantos “cientistas midiáticos” são conhecidos hoje em dia no Brasil? Talvez a professora Mayana Zatz, da USP. Ela tem um blog na VEJA, chamado Genética, no qual discute vários assuntos relacionados a este tema. Mas a postagem que causou a maior discussão não foi nenhuma destas, e sim sobre “O caso de Sean e o direito de decidir”. Mayana foi agredida verbalmente por muitos leitores de seu blog, por emitir sua opinião sobre o caso do direito de guarda do menino Sean Goldman. Foi uma das postagens que menos falou sobre ciência.
Pensando sobre este assunto, cheguei à conclusão que ainda existe um longo caminho a ser feito, de aproximação dos cientistas da mídia e vice-versa, no sentido de se desmistificar os cientistas e o conhecimento científico como sendo assunto somente de especialistas. A divulgação científica de qualidade é extremamente importante, e a sociedade quer saber, quer conhecer, quer aprender. Levando em conta que a sociedade paga pela quase totalidade do desenvolvimento científico no Brasil, a divulgação científica de qualidade deixa de ser uma opção e passa a ser uma obrigação Cabe a nós, cientistas, e aos meios de comunicação, juntos trabalhar para mostrar que a ciência e os cientistas não são seres de outro planeta. A necessidade de cientistas midiáticos pode ser real – mas de uma mídia responsável, mais p
róxima dos cientistas e da ciência, também é.
Em tempo: sobre este mesmo assunto, o artigo
“A Arte do Desencontro” de Mariluce Moura, editora-chefe da
revista Pesquisa FAPESP, foi publicado no último número (junho de 2010)
desta revista.
103 (melhores?) livros de ciência

20,000 Microbes Under the Sea, Robert Kunzig
A Gene for Nothing- A Natural History of Peace, Robert Sapolsky
A Leg to Stand On, Oliver Sacks (2 indicações)
A Linguistic Big Bang, Lawrence Osborne
A Mathematician’s Apology, G.H. Hardy (2 indicações)
A Medical Quest, Masha Gessen
Altered Oceans, Kenneth Weiss e Usha Macfarling
Anything and everything, Michio Kaku, Steven Pinker e James Lovelock.
Chaos: Making a New Science, James Gleick
Consciousness Explained, Dan Dennett (2 indicações)
Cosm, Gregory Benford
Cosmos, Carl Sagan
De Humani Corporis Fabrica (On the Fabric of the Human Body), Andreas Vesalius (1543)
De Revolutionibus Orbium Coelestium (On the Revolutions of Heavenly Spheres), Nicolaus Copernicus (1543)
Dialogue Concerning the Two Chief World Systems, Galileo Galilei (1632)
Disturbing the Universe, Freeman Dyson
Does God Play Dice, Ian Stewart
Faber Book of Science, John Carey
Flatland, Edwin Abbott.
Foundation, Isaac Asimov Gaia, James Lovelock
Genethics, David Suzuki & Peter Knudston
Genome, Matt Ridley
George Divoky’s Planet, Darcy Frey
Gödel, Escher, Bach, Douglas Hofstadter (3 indicações)
Gorillas in the Mist, Dian Fossey (1983)
Guns, Germs and Steel, Jared Diamond.
How to Build a Time Machine, Paul Davies (2 indicações)
Infinity Plus One, Polly Shulman
Invention, Norbert Wiener (2 indicações)
Is Evolution Wrong?, David Quammen
Kon Tiki, Thor Heyerdahl and F.H.Lyon.
Last and First Men, Olaf Stapledon
Late Night Thoughts on Listening to Mahler’s Ninth Symphony, Lewis Thomas
Mason & Dixon, Thomas Pynchon (2 indicações)
Mathematics, Form and Function, Saunders Mac Lane (2 indicações)
Micrographia, Robert Hooke (1665)
Monkeyluv, Robert Sapolsky
Moonshine and Glue: A Thirteen-Unit Guide to the Extreme Edge of Astrophysics, Oliver Morton
Mother Earth Mother Board, Neal Stephenson
Mother Nature: Natural Selection & the Female of the Species, Sarah Blaffer Hardy.
Nothing in Biology Makes Sense Except in the Light of Evolution, Theodosius Dobzhansky
Of a Fire on the Moon, Norman Mailer.
On Being the Right Size, J.B.S. Haldane
On Growth and Form, D’Arcy Wentworth Thompson (2 indicações)
One Two Three . . . Infinity, George Gamow (1947)
Phantoms in the Brain, V.S.Ramachandran
Philosophiae Naturalis Principia Mathematica (Mathematical Principles of Natural Philosophy), Isaac Newton (1687)
Physica (Physics), Aristotle (circa 330 B.C.)
Pluto’s Republic, Peter Medawar (2 indicações)
Relativity: The Special and General Theory, Albert Einstein (1916)
Saving Us From Darwin, Frederick Crews
Science and Civilisation in China, Joseph Needham
Science and Music, Sir James Jeans.
Sexual Behavior in the Human Male, Alfred C. Kinsey et al. (1948)
Shadows of the Mind, Roger Penrose (2 indicações)
Silent Spring, Rachel Carson (1962) (2 indicações)
Something New Under the Sun: An Environmental History of the Twentieth Century, John McNeill.
Surely You’re Joking, Mr. Feynman!, Richard Feynman (4 indicações)
The Abyss, Oliver Sacks
The Art of the Soluble, Peter Medawar
The Beak of the Finch, Jonathan Weiner
The Black Cloud, Fred Hoyle
The Blank Slate, Steven Pinker (2 indicações)
The Brain on the Stand, Jeffrey Rosen
The Chimpanzees of Gombe, Jane Goodall
The Cosmic Connection, Carl Sagan (1973)
The Curse of Qwerty, Jared Diamond
The Double Helix, James D. Watson (1968) (4 indicações)
The Existential Pleasure of Engineering, Samuel C. Florman
The Extended Phenotype, Richard Dawkins.
The Feynman Lectures on Physics, Richard P. Feynman, Robert B. Leighton, e Matthew Sands (1963)
The First Three Minutes, Steven Weinberg (1977)
The Insect Societies, Edward O. Wilson (1971)
The Itch, Atul Gawande
The Journals of Lewis and Clark, Meriwether Lewis and William Clark (1814)
The Language Instinct, Steven Pinker.
The Life of Galileo, Bertold Brecht (2 indicações)
The Making of the Atomic Bomb, Richard Rhodes
The Man Who Mistook His Wife for a Hat and Other Clinical Tales, Oliver Sacks (1985) (3 indicações)
The Microbe Hunters, Paul de Kruif (2 indicações)
The Mismeasure of Man, Stephen Jay Gould (1981)
The Origin of Species, Charles Darwin (1859) (2 indicações)
The Panda’s Thumb, Stephen Jay Gould
The Prehistory of the Mind, Steven Mithen
The Road to Reality: A Complete Guide to the Laws of the Universe, Roger Penrose
The Selfish Gene, Richard Dawkins (1976)
The Stars My Destination, Alfred Bester.
The Strategy of Conflict, Thomas Schelling (2 indicações)
The Velvet Claw, David Macdonald.
The Voyage of the Beagle, Charles Darwin (1845) (3 indicações)
Under a Lucky Star, Roy Chapman Andrews (1943)
What if It’s All Been a Big Fat Lie?, Gary Taubes
What Is Life?, Erwin Schrödinger (1944)
arities – Landmarks on the Pathways of Life”, por Christian de Duve, e “The Origins of Life” por John Maynard Smith e Eörs Szathmáry. Ambos apresentam excelentes argumentos sobre o surgimento e o desenvolvimento da vida no planeta Terra.

Uma amiga me disse certa vez que sua angústia era saber que iria morrer sem conseguir ler tudo o que gostaria. Fiquei surpreso com a angústia dela, e pensei se esta seria uma de minhas angústias também. Não sei. Existe um livrinho chamado “Sabedoria Chinesa”, organizado por Dagmar Von Berg (Editora Pensamento, 1984), de frases ou ditados da filosofia chinesa. Uma das frases diz o seguinte: “Um livro é como um jardim de bolso”.
Ruminantes, plantas e efeito estufa
Consequentemente, o investimento em pesquisa para se minimizar a produção de metano por ruminantes têm sido significativo. Foram desenvolvidos vários aditivos químicos para serem incorporados ao alimento do gado para se diminuir a produção de metano. Porém, vários destes aditivos apresentam certo grau de toxicidade, e atuam apenas de maneira parcial; muitos não são completamente degradados no trato digestivo destes animais, e podem gerar resíduos químicos indesejáveis; finalmente, alguns destes aditivos são antibióticos para reduzir a flora de bactérias que produzem metano (bactérias metanogênicas), e estas bactérias podem adquirir resistência a estes antibióticos. Ou seja, tais aditivos químicos estão longe de ser a solução ideal para a redução da emissão de gases metano por ruminantes.
Uma alternativa é o uso de plantas que apresentam substâncias químicas que inibem bactérias metanogênicas, ou ainda a produção de metano por estas bactérias. Estas plantas apresentam substâncias químicas conhecidas como produtos do metabolismo secundário (produtos naturais), que interagem com os micro-organismos da flora dos ruminantes e podem contribuir significativamente para a redução da produção de metano.

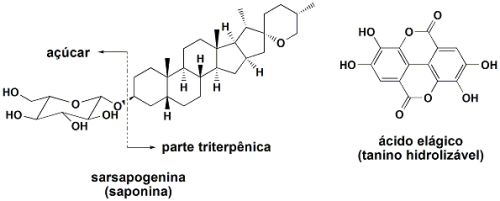
Saponinas
Saponinas são derivados de triterpenos glicosilados. Triterpenos são substâncias com 30 átomos de carbono, ou menos (porque alguns átomos de carbono são eliminados nos caminhos bioquímicos que levam à formação dos “triterpenos degradados”). Por exemplo, os esteróis e esteróides se originam de uma cadeia de 30 átomos de carbono, a qual pode ser diminuída a até 25, dependendo de como estas substâncias são formadas. As saponinas são triterpenos ou esteróis modificados, ligados a uma ou mais de uma molécula de açúcares (existem 8 açúcares naturais com seis átomos de carbono, as hexoses). O nome “saponina” se origina do fato que muitas destas substâncias atuam como detergentes, até mesmo produzindo espuma em meio aquoso. As saponinas têm ampla distribuição no reino vegetal.
Observou-se que grandes concentrações de saponinas causam efeito direto sobre as bactérias metanogênicas, fazendo com que estas diminuam a produção de metano. Algumas saponinas diminuem a atividade de genes reguladores da produção de metano, ou a taxa de produção de metano em células metanogênicas. Alguns protozoários presentes no trato digestivo de ruminantes também podem produzir metano, e algumas saponinas apresentam ação anti-parasítica contra estes protozoários, ligando-se às suas membranas celulares.
A sarsaponina, presente no amido da batata, é a saponina mais efetiva como redutora da metanogênese.
O mais interessante é que a presença de saponinas em plantas adicionadas ao alimento dos ruminantes não diminui a palatabilidade dos alimentos, ou a sua assimilação. Ou seja, a utilização de plantas com saponinas não diminui a qualidade nutricional dos alimentos oferecidos para ruminantes. Na verdade, em alguns casos a presença de saponinas nos alimentos dos ruminantes parece estimular o apetite destes animais. A eventual perda de “digestibilidade” devido à presença de saponinas é que estas podem apresentar atividade anti-bacteriana ou anti-fúngica contra micro-organismos celulolíticos presentes no trato digestivo dos ruminantes. Estes micro-organismos têm enzimas que degradam a celulose presente nos vegetais dos quais os animais se alimentam. Este é o único impedimento observado no caso de se utilizar plantas com saponinas como aditivo aos alimentos de ruminantes.
Taninos
Taninos são polímeros polifenólicos hidrossolúveis (solúveis em água), alguns de alto peso molecular, que podem se agregar com proteínas. Os taninos estão presentes em muitas plantas utilizadas na alimentação de ruminantes, como gramíneas, legumes, frutas, cereais e grãos.
Não se tem informações precisas sobre os efeitos de taninos sobre bactérias metanogênicas. A ação anti-metanogênica dos taninos pode estar relacionada à inativação de proteínas funcionais (enzimas). Além disso, a ação dos taninos depende do tipo de tanino presente na planta incluída na dieta dos ruminantes, bem como da quantidade em que estão presentes nestas plantas. Provavelmente sua ação também inclui a inibição do crescimento de bactérias metanogênicas, bem como uma possível ação bactericida.
Vários tipos de plantas utilizadas na alimentação de ruminantes apresentam taninos que promovem a diminuição da produção de metano pelas bactérias metanogênicas. Também alguns frutos utilizados para o mesmo fim podem reduzir a produção de metano. O problema é que várias bactérias desenvolvem a capacidade de metabolizar taninos, e estes perdem sua eficácia após algum tempo de uso na ração de ruminantes. Bactérias metanogênicas outras do que aquelas encontradas nos animais mostraram ser suscetíveis a taninos até por 2 meses, no máximo.
Outro problema associado à inclusão de plantas com taninos na dieta de ruminantes é que os taninos afetam a palatabilidade do alimento, bem como da digestibilidade do alimento. Taninos podem ser incluídos em até no máximo 5% do total dos aditivos adicionados à dieta dos animais. A ação antibiótica dos taninos pode comprometer a flora digestiva dos ruminantes, prejudicando sua capacidade de digestão. Logo, o uso de taninos para reduzir a metanogênese apresenta eficácia limitada.
Óleos essenciais
Os óleos essenciais vegetais constituem a parte volátil da composição das plantas. São substâncias químicas de baixo peso molecular, frequentemente utilizadas como aromatizantes e flavorizantes (para melhorar o aroma e o sabor de alimentos). Os dois principais grupos de constituintes dos óleos essenciais são os terpenos e os derivados fenilpropanoídicos. Dentre os primeiros se incluem os monoterpenos (com 10 átomos de carbono) e os sesquiterpenos (com 15 átomos de carbono). Já os derivados fenilpropanoídicos são formados por 9 átomos de carbono.
Os terpenos são extremamente diversificados, e estão presentes em todas as plantas. Já os derivados fenilpropanoídicos são menos abundantes e menos diversificados do que os terpenos, mas algumas plantas (como o cravo e a canela) apresentam derivados fenilpropanoídicos em grandes quantidades. Os óleos essenciais podem ser obtidos de diferentes partes das plantas – flores, pétalas, folhas, frutos, raízes e casca de árvores.
Observou-se que os óleos essenciais aumentam a diversidade de algumas bactérias metanogênicas (bactérias do tipo Archea). Porém, no caso das bactérias metanogênicas comuns, o efeito dos óleos essenciais é de diminuir a produção de metano. Um dos primeiros estudos realizados com óleos essenciais para a diminui
Antibióticos em falta
A relação numérica entre humanos e bactérias é de 1 para 1 bilhão de trilhões, sendo que a taxa de reprodução das bactérias é de cerca de 500.000 vezes mais rápida que dos humanos. Ou seja, o mundo é literalmente dominado por bactérias, por mais desinteligente que tal design possa parecer. Além do mais, algumas bactérias são especialmente nocivas. A espécie Staphylococcus aureus resistente à meticilina, por exemplo (MRSA methicillin-resistant Staphylococcus aureus), é a responsável direta por 100.000 mortes/ano nos EUA. Outras espécies particularmente perniciosas por serem resistentes a muitos fármacos são Pseudomonas, Acinetobacter, Klebsiella. O tratamento de infecções causadas por estas bactérias patogênicas é bastante limitado, pois apenas um número pequeno de antibióticos é efetivo para eliminá-las.
Como a descoberta de novos antibióticos têm sido cada vez mais difícil, médicos estão fazendo uso cada vez mais freqüente de antibióticos de segunda e terceira linha, que são mais custosos, menos efetivos e mais tóxicos. Para se ter uma idéia, o tratamento de uma infecção causada por uma bactéria resistente custa o dobro do tratamento de uma infecção comum, e o risco de morte por tal infecção é duas vezes maior. Além disso, a disseminação de bactérias resistentes em ambientes hospitalares é muito mais fácil se medidas de higiene extremamente eficazes não são tomadas, como desinfecção de mãos e pés, além dos sistemas de ventilação, e até mesmo de cateteres. Estima-se que 80% das infecções hospitalares poderiam ser prevenidas apenas por melhores medidas de assepsia.
Mesmo adotando-se as melhores práticas hospitalares, as bactérias resistentes não irão desaparecer, pelo simples fato destas estarem presentes no ambiente há muitos milhões de anos. Bactérias e fungos estão em contínua competição entre si e com outros seres vivos, como insetos, plantas e outros animais, e por isso desenvolveram mecanismos de resistência muito eficazes para poder competir no ambiente natural. Para se ter uma idéia, um estudo realizado por pesquisadores da McMaster University demonstrou que um terço de 500 bactérias isoladas de solos são resistentes a 1/3 de todos os antibióticos aos quais foram expostas. Outros estudos demonstraram que a resistência de bactérias ao antibiótico eritromicina surgiu há aproximadamente 880 milhões de anos, e a resistência à penicilina (produzida por fungos) deve ter surgido há cerca de 2 bilhões de anos, sendo até mesmo anterior à diferenciação entre bactérias Gram-positivas e Gram-negativas.
Algumas bactérias apresentam a capacidade de eliminar os antibióticos de sua célula antes mesmo destes fazerem efeito, uma capacidade chamada de “mecanismos de efluxo bacteriano”. As bactérias Gram-negativas apresentam paredes celulares que bloqueiam a entrada de antibióticos. Pseudomonas, por exemplo, apresenta as duas características, e pode ser extremamente perniciosa. Outras bactérias desenvolveram enzimas para neutralizar ou destruir antibióticos, como as penicilinas, chamadas de ß-lactamases.
Outra característica única das bactérias é a sua capacidade de trocar genes com outras baterias, como nós trocamos figurinhas de álbuns da copa do mundo. Ao trocar genes as bactérias podem compartilhar genes de resistência a diferentes antibióticos. Desta maneira, a utilização abusiva e a má utilização de antibióticos, que é muito freqüente, faz com que as bactérias passem a adquirir resistência a cada vez um número maior de antibióticos. Consequentemente, a disponibilidade de antibióticos para o tratamento de infecções está cada vez mais restrita.
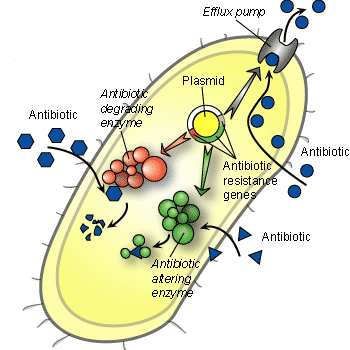
Na verdade, a “era de ouro” de descoberta dos antibióticos foi entre 1945 e 1960, quando a grande maioria dos antibióticos que hoje são utilizados foi isolada de bactérias e fungos. Desde então, muito poucos novos antibióticos foram descobertos – a grande maioria resulta de variações dos antibióticos descobertos há mais de 50 anos. Para que se possam desenvolver novos antibióticos, mais eficazes, é necessário que se descubram classes novas destes fármacos, que atuem de maneira diferente dos antibióticos já conhecidos.
Embora o seqüenciamento de genomas na década de 90 tenha causado grande excitação na indústria farmacêutica, nenhum novo antibiótico resultou das pesquisas genômicas. Entre 1995 e 2001 a indústria farmacêutica GlaxoSmithKline testou quase 500.000 de substâncias diferentes em 300 diferentes alvos terapêuticos bacterianos, sem um único sucesso. Como resultado, as indústrias farmacêuticas passaram a deixar de lado a pesquisa por novos antibióticos. Preferiram investir esforços na descoberta de fármacos mais fáceis de serem desenvolvidos, e bem mais rentáveis, como “remédios” para impotência e calvície.
As causas das falhas de novos antibióticos foram muitas. Vários antibióticos apresentaram espectro de ação muito mais restrito do que o desejado para serem clinicamente válidos (e de interesse comercial também). Outros apresentaram altos níveis de toxicidade, além de não apresentarem propriedades adequadaas para serem utilizados como medicamentos (baixa solubilidade em plasma, baixa capacidade de atravessar membranas, baixa absorção, etc.). Além de descobrir novos antibióticos eficazes, o maior problema foi de introduzir um novo fármaco no mercado que pudesse ser utilizado por um tempo razoável. Como os agentes patogênicos podem adquirir resistência relativamente rápido, a eficácia dos antibióticos fica limitada a poucos anos. A combinação de todos estes fatores torna excessivamente alto o investimento na descoberta de novos antibióticos. Além das exigências de agências sanitárias, como o FDA (Food and Drug Adminsitration, dos EUA), serem muito rigorosas para a introdução de novos antibióticos no mercado.
Desta maneira, os médicos estão passando a adotar novas formas de tratamento, principalmente baseadas na prevenção. Mesmo assim, existem situações em que antibióticos são imprescindíveis. Caso não se redobrem esforços para se descobrir novas classes de antibióticos, muito em breve estaremos em uma sinuca de bico.
Shekhar, C. (2010). Bacteria: Drug Resistance Spreads, but Few New Drugs Emerge Chemistry & Biology, 17 (5), 413-414 DOI: 10.1016/j.chembiol.2010.05.
006
Homenagem ao Dia Mundial do Meio Ambiente de 2010
Como os fatos mostram que o ambiente em que vivemos não vai muito bem, decidi fazer uma homenagem poética e musical a este dia tão especial.
Primeiro com uma música de Edu Lobo, Augusto Boal e Gianfrancesco Guarnieri, gravada originalmente em 1967. A música foi composta para a peça de teatro “Arena conta Zumbi”, e se insere no momento em que negros estão se articulando para criar um quilombo, mas alguns estão em dúvida. Zumbi então tenta convencer os indecisos, cantando a música. É meio anti-ambientalista, pois os personagens querem “bichos prá comer”, e meio machista também. Mas, há que se considerar a época de sua criação. Escute a música “clicando” no botão “>” a seguir (agradecimentos especiais ao Igor Santos que me ensinou como inserir o arquivo mp3 da música “embolada”). Vejam também uma parte da encenação da peça “Arena conta Zumbi” por estudantes do Colégio Cenecista Dr. José Ferreira, justamente no trecho em que a música é cantada: http://www.joseferreira.com.br/tvzezao.php?v=17825&c= . Emocionante! (valeu, Sibele, pela dica).
De toda forma e qualidade tem
oi tem pindoba, embiriba e sapucaia
tem titara, catulé, ouricurí
tem sucupira, sapucais, putumujú
teu pau-de-santo, tem pau d’arco, tem tatajubá
sapucarana, canzenzé, maçaranduba
tem louro paraíba e tem pininga (bis)
Pare meu irmão
de falar em tanta mata
com tanta planta eu não sei o que fazer
mas diga lá se tem bicho pra comer
se tem bicho pra comer, se tem bicho pra comer.
De toda forma e qualidade tem,
onça pintada, sussuarana e maracajá
E tem guará, jaguatirica e guaxinim
e tem tatu, tatu-peba, tatu-bola
tem preguiça, tem quatí, tamanduá.
E coelho que tem, tem, tem
queixada que tem, tem, tem
caititú oi tem também
oi diz que tem, tem
oi diz que tem, tem (bis).
Pare meu irmão
de falar em tanta fera
com tanto bicho eu não sei o que fazer
ah, um bichinho pra comer
eu só quisera
com tanto assim eles vão é me comer (bis).
Mas tem os peixes que ainda não falei
de toda forma e qualidade tem
oi tem traíra, tem cará e jundiá
e tem caborge, tem piaba e carapó
e pitú e caranguejo e aruá (repete mais 3 vezes).
Mas também tem cobra
que é um nunca se acabar
tem jacaré, cobra-rainha e tem muçu
tem caninana, tem jibóia e tem jericoá
tem jararaca, cascavel, surucucú
e papa-ovo e cobra verde assim não dá (bis).
Mas tem sabiá, tem canário e curió
tem passarinho tão bom de se olhar
papa-capim, cardeal e arumará
e tem xexéu, guriatã e tem brejá
E se quiser comer galinha
tem de todas pra fartar
tem pomba de tres côcos, tem pato mergulhão
aracuâ, jaçanã e tem carão
juriti e cardigueira e paturí (repete mais 3 vezes).
Mas e nessa abençoada região
será que tem o que faz falta na verdade?
O que é, o que é, o que é ?
O que é, o que é, o que é ?
Me diga meu irmão
se nessa grande mata (BIS)
é possível, é possível ter mulher.
Aí está uma coisa que não…
Aí está uma coisa que não…
Pois sendo assim eu prefiro o cativeiro.
Pois sendo assim eu prefiro o cativeiro.
Meu irmão tá com toda razão !
Meu irmão tá com toda razão !
A segunda é do Chico Buarque de Hollanda e do Francis Hime, e foi composta para o filme “A Noiva da Cidade” de Alex Vianni.
Ei, pintassilgo
Oi, pintaroxo
Melro, uirapuru
Ai, chega-e-vira
Engole-vento
Saíra, inhambu
Foge asa-branca
Vai, patativa
Tordo, tuju, tuim
Xô, tié-sangue
Xô, tié-fogo
Xô, rouxinol sem fim
Some, coleiro
Anda, trigueiro
Te esconde colibri
Voa, macuco
Voa, viúva
Utiariti
Bico calado
Toma cuidado
Que o homem vem aí
O homem vem aí
O homem vem aí.
Ei, quero-quero
Oi, tico-tico
Anum, pardal, chapim
Xô, cotovia
Xô, ave-fria
Xô, pescador-martim
Some, rolinha
Anda, andorinha
Te esconde, bem-te-vi
Voa, bicudo
Voa, sanhaço
Vai, juriti
Bico calado
Muito cuidado
Que o homem vem aí
O homem vem aí
O homem vem aí
Borzeguim, deixa as fraldas ao vento
E vem dançar
E vem dançar
Hoje é sexta-feira de manhã
Hoje é sexta-feira
Deixa o mato crescer em paz
Deixa o mato crescer
Deixa o mato
Não quero fogo, quero água
(deixa o mato crescer em paz)
Não quero fogo, quero água
(deixa o mato crescer)
Hoje é sexta-feira da paixão sexta-feira santa
Todo dia é dia de perdão
Todo dia é dia santo
Todo santo dia
Ah, e vem João e vem Maria
Todo dia é dia de folia
Ah, e vem João e vem Maria
Todo dia é dia
O chão no chão
O pé na pedra
O pé no céu
Deixa o tatu-bola no lugar
Deixa a capivara atravessar
Deixa a anta cruzar o ribeirão
Deixa o índio vivo no sertão
Deixa o índio vivo nu
Deixa o índio vivo
Deixa o índio
Deixa, deixa
Escuta o mato crescendo em paz
Escuta o mato crescendo
Escuta o mato
Escuta
Escuta o vento cantando no arvoredo
Pas
sarim passarão no passaredo
Deixa a índia criar seu curumim
Vá embora daqui coisa ruim
Some logo
Vá embora
Em nome de Deus é fruta do mato
Borzeguim deixa as fraldas ao vento
E vem dançar
E vem dançar
O jacu já tá velho na fruteira
O lagarto teiú tá na soleira
Uirassu foi rever a cordilheira
Gavião grande é bicho sem fronteira
Cutucurim
Gavião-zão
Gavião-ão
Caapora do mato é capitão
Ele é dono da mata e do sertão
Caapora do mato é guardião
É vigia da mata e do sertão
(Yauaretê, Jaguaretê)
Deixa a onça viva na floresta
Deixa o peixe n’água que é uma festa
Deixa o índio vivo
Deixa o índio
Deixa
Deixa
Dizem que o sertão vai virar mar
Diz que o mar vai virar sertão
Deixa o índio
Dizem que o mar vai virar sertão
Diz que o sertão vai virar mar
Deixa o índio
Deixa
Deixa
