
Inferir o comportamento e a função de organismos extintos por meio dos registros fósseis é sempre um assunto polêmico. Alguns paleontólogos afirmam que isso deveria ser evitado, já que as hipóteses propostas nunca poderiam ser testadas, porém outros consideram essa uma das tarefas primordiais da paleontologia…
Nesse post dos colecionadores, inspirada pelo artigo do Prof. Michael J. Benton da Universidade de Bristol, Inglaterra, publicado em março deste ano na revista PLoS Biology, pretendo abordar um assunto um tanto controverso e espero que isso inspire alguns dos leitores a fazerem comentários. A inferência da função e comportamento de animais extintos por meio do registro fóssil. Ciência ou não? Qual a importância de tentar compreender detalhes da vida de organismos extintos?
Benton começa muito bem seu artigo, relembrando o fundador da anatomia comparada, Georges Cuvier (1769-1832). Georges Cuvier, naturalista francês, que foi ilustre titular da cadeira de Anatomia Animal do Museu de História Natural de Paris no início do século XIX, foi quem instituiu a observação de que existe um padrão comum entre os esqueletos de vertebrados, tanto os fósseis quanto os viventes. E foi baseado nisso que ele propôs a possibilidade de se reconstruir a anatomia de um animal extinto a partir até mesmo de registros fósseis incompletos.
Fundamentado nesse seu princípio básico da anatomia e levando em consideração todas as suas observações de organismos vindos de vários cantos do planeta, Cuvier argumentou que o esqueleto de um animal, existente ou extinto, guarda pistas inequívocas sobre sua função e comportamento. Assim, enxergou que sua missão seria estabelecer algumas regras para a anatomia comparada que permitissem aos paleontólogos realizar certas afirmações sobre animais fósseis com clareza e confiança.
Isso se tornou um princípio-chave na atualidade, e é o que chamamos de “reconstrução baseada na evidência” (i.e. dentes afiados indicam uma dieta carnívora, porque esse é um padrão típico observado nos organismos) diferente da pura especulação (i.e. “este dinossauro era rosa porque eu acho que ele era”).
Forma, função e comportamento
Pode-se assumir que estruturas biológicas são adaptadas de alguma forma e que elas evoluíram de maneira razoavelmente eficiente para realizar uma determinada função. Assim seria a tromba de um elefante: um órgão que evoluiu e atua como ferramenta muito competente de sucção e preênsil, permitindo ao grande animal alcançar o solo para buscar água e comida. A partir de observações semelhantes, Cuvier entendeu que a forma de uma estrutura biológica deveria então refletir a sua função. Cuvier, no entanto, interpretou isto como sendo evidência da existência de algum tipo design inteligente e não como produto da evolução. É útil lembrar-se desse detalhe histórico, porque cabe colocar aqui, então, um alerta. É necessário cautela nas interpretações: nem tudo é resultado de uma adaptação e nem toda ‘adaptação’ é perfeita – A evolução trabalha como um sucateiro, não há uma escolha inteligente ou uma entidade por trás disso tudo, a evolução trabalha com os materiais que ela tem nas mãos.
Os fósseis como poderosas fontes de informação
Fósseis podem fornecer uma grande quantidade de informações:
1. Anatômicas gerais dos organismos: O formato do esqueleto ou de outras partes biomineralizadas , que são os elementos mais facilmente preservadas no registro fóssil (i.e. o esqueleto reforçado dos artrópodes fósseis, que revela o formato de seu corpo e o número dos apêndices, a natureza de cada articulação e até detalhes das peças bucais e outras estruturas relacionadas com a alimentação e locomoção, as conchas de moluscos, dentes, escamas e ossos nos vertebrados).
2. Anatômicas mais especializadas: Como evidências de cicatrizes de inserção muscular no esqueleto de vertebrados ou nas conchas de moluscos (i.e. impressões e alguns processos indicam de que maneira o músculo estava inserido ou qual era o seu tamanho, o que permite calcular a seu volume e potência) ou outras biomecânicas, como o cálculo do máximo rotacional de cada articulação dos ossos, inferido com base no formato das extremidades dos mesmos. Esse último pode ajudar a resolver questões interessantes sobre a postura e locomoção de animais extintos (i.e. a postura de caminhar nos pterossauros seria parasagital (com os membros abaixo do corpo) ou esparramada como a de um cowboy? Os fósseis demonstraram que a última seria o caso).
Mas não somente.Fósseis excepcionalmente bem preservados podem revelar muitos detalhes adicionais:

Anchiornis huxleyi
- Como os contornos de partes moles dos organismos (i.e. dos tentáculos de beleminitas ou amonitas, de órgãos internos, pele e músculos de dinossauros, dos olhos compostos de trilobitas, etc.)
- A presença de pêlos, penas ou outras estruturas semelhantes – em mamíferos, pterossauros, dinossauros ou aves.
- Ou mesmo detalhes inesperados de estruturas moleculares (i.e. A preservação de osteócitos, de melanossomos, ou mesmo de boas porções de DNA, como nos mamutes congelados da Sibéria).
- Existem ainda os fósseis excepcionais que preservam evidências de interações, mas vamos nos referir a eles mais detalhadamente adiante.
Abordagens para se extrair ao máximo as informações dos fósseis: A Função e Comportamento!
Existem três abordagens para inferir função e comportamento por meio de fósseis – 1) evidências empíricas, 2) comparação com análogos modernos e 3) a modelagem biomecânica. Vamos considerar as três:
1) Evidências empíricas
Paleontólogos são inquisitivos por natureza, e eles reúnem evidências de todos os tipos para testar suas hipóteses. Evidências sobre o estilo de vida de um organismo extinto podem estar contidas 1) na rocha matriz, 2) na assembléia fossilífera associada (os outros organismos fósseis encontrados no mesmo contexto ou mesmo icnofósseis associados) e 3) em características particulares do próprio fóssil em questão.
As rochas podem nos dar evidências claras sobre paleoclimas e os fósseis associados podem indicar potenciais interações (predadores ou presas, etc). Já os icnofósseis, como pegadas e escavações, podem algumas vezes ser conectados aos seus produtores, e no caso de coprólitos (fezes fósseis), podem ser vasculhados para se determinar constituintes-chave da dieta de animais extintos.
Um famoso coprólito de 44 cm associado à Tyrannosaurus rex contém restos pulverizados de ossos de dinossauros Ornitischia que foram expostos à ação de ácidos estomacais, mas não foram inteiramente destruídos. Isso sugere um transito relativamente rápido do material alimentar pelo trato digestivo desses animais.
Quanto a características específicas do fóssil em questão, podemos, por exemplo, considerar detalhes dos dentes de animais pré-históricos para determinar a sua dieta.
Um estudo detalhado dos dentes de mamíferos fósseis pode até mesmo indicar que tipos de plantas eles utilizavam preferencialmente como recursos, baseando-se em micro-abrasões observáveis somente com o uso de microscópios eletrônicos. Ainda podem ser feitas análises das assinaturas isotópicas dos dentes, que revelam preferências por plantas C3 ou C4 na dieta dos animais, reflexos do paleoambiente no qual esses organismos estavam inseridos.
Algumas vezes um organismo pode ainda ser preservado de uma forma excepcional: capturado em um flagrante – flagras de interações: Alimentando-se, por exemplo, ou mesmo engajado em um confronto. Já fizemos referência a esses tipos de fósseis em um tópico mais antigo desse blog. Tratava-se particularmente do relato de um fóssil indiano descrito por Jeff Wilson e colaboradores. Sanajeh indicus, uma cobra preservada quase completa em pleno ataque a um ninho de dinossauros saurópodes.

Fóssil de Velociraptor e Protoceratops engajados em uma luta mortal.

O Flagrante fóssil de um Peixe abocanhando o outro. O resultado foi mortal.

Fóssil e reconstruções de Sanajeh indicus.
Na ocasião, Wilson e colaboradores utilizaram todos os três métodos comentados aqui (observações empíricas do espécime, combinada com comparações com análogos modernos e reconstrução biomecânica) para oferecer uma imagem do que retrataria aquele fóssil. Os autores incorporaram pesquisas museológicas, pesquisas de campo, estratigrafia e sedimentologia, histologia, embriologia e até mesmo análogos modernos para interpretar os detalhes de Sanajeh. Chegaram à conclusão de que aquele animal estaria espreitando o ninho de saurópodes à espera dos recém-nascidos para apanhá-los. É claro que não podemos ter completa certeza disso até que novos espécimes sejam encontrados com evidências de ossos dos pequenos bebês no que seria a região estomacal da cobra. MAS, em todos os casos, essas não são meras observações ou descrições óbvias de dados. São sim, hipóteses, como quaisquer outras, com suas referências e métodos, e sujeitas à refutação a qualquer momento.
O exemplo de que uma mandíbula que possua dentes mamalianos possa informar a um paleontólogo que aquele material pertença, vamos dizer, à uma toupeira marsupial de uma determinada família, e que devido aos seus dentes serem formato de agulha, ela deveria alimentar-se de insetos, está inteiramente dentro do modelo hipotético-dedutivo.
Assim como a asserção clássica de que “todos os cisnes são brancos” foi refutada pela descoberta do cisne negro Tasmaniano, toda asserção ou alegação feita por um paleontólogo está aberta a uma inspeção mais detalhada e refutação por novas evidências.
Comparação com análogos modernos
É provável que a função e o comportamento de um morcego fóssil possam ser inferidas com base em comparações com morcegos modernos, mas deveria um dinossauro ser comparado com seus parentes viventes (crocodilos e aves) ou animais com função ecológica aparentemente similar (elefantes ou rinocerontes)? A filogenia triunfa sobre similaridades gerais?
Talvez seja inútil comparar um Diplodocus com um pardal – seu tamanho corporal, morfologia e presumíveis modos de vida são completamente diferentes. Mas algo informativo vem sim da filogenia. De um modo ou de outro, a parcimônia permite aos paleobiólogos inferir sobre a presença de alguns tipos de caracteres e comportamentos. O desenvolvimento desse princípio da parcimônia é o que chamamos de “extant phylogenetic bracket” ou EPB.
De acordo com esse princípio, de uma maneira simples: poderíamos dizer que Tyrannosaurus rex presumivelmente teria os olhos com certas características, porque seus parentes viventes – aves e crocodilos – compartilham muitos caracteres em comum nos seus olhos. Outro exemplo, talvez um pouco mais impressionante, é a predição de que ovos fósseis serão um dia encontrados em estratos do período Carbonífero (360-280 m.a.). A razão principal para isso é que todos os amniotas viventes (i.e. répteis, aves e mamíferos) depositam ovos com casca, com exceção dos mamíferos que substituíram essa forma reprodutiva para dar a luz diretamente aos seus filhotes e também algumas espécies de cobras e lagartos. Então, recapitulando: considerando a validez dessa observação para a atualidade, espera-se que o primeiro amniota no Carbonífero, há mais de 300 milhões de anos atrás, presumivelmente depositava ovos com casca, mesmo que o mais antigo registro para ovos fósseis seja datado do Triássico, 100 milhões de anos depois. Espera-se encontrar registros mais antigos.
A Parcimonia e o EPB são agora largamente utilizados nos discursos sobre os dinossauros com penas e as aves do Grupo Jehol da China (Cretácio Inferior, 131-120 m.a.). Quando espécimes do pequeno dinossauro terópode Sinosauropteryx foram anunciados, com a presença de filamentos simples análogos a penas cobrindo quase todo seu corpo, paleontólogos logo olharam para as árvores filogenéticas e concluíram que essa descoberta lançava a origem das penas bem para trás no tempo, para a base do Jurássico Médio, 175 milhões de anos atrás. Isso porque Sinosauropteryx é um coelurossauro basal, e os primeiros coelurossauros conhecidos pertencem ao início do Jurássico Médio. A suposição mais parcimoniosa é que todos os coelurossauros possuíam algum tipo de pena desde o início de sua evolução.
Note que os filamentos de Sinosauropteryx ainda encontram-se em debate. Alguns pesquisadores argumentam que eles não seriam penas, mas sim tecido conectivo. De qualquer forma, penas tem sido reportadas para quase todas as linhagens de coelurossauros encontradas, e, portanto, sua origem estaria de alguma forma profundamente enraizada na filogenia dos dinossauros terópodes. Conhecendo o arranjo das penas e agora até mesmo o seu padrão de coloração, os paleobiólogos muito em breve poderão especular calmamente e racionalmente, sobre se certos dinossauros utilizavam suas penas para camuflagem, para emitir sinais de alerta, como display sexual, para estabelecer ordens sociais ou ainda outros comportamentos e funções.
Modelagem biomecânica
Modelos biomecânicos, combinados com considerações de análogos modernos, fornecem um poderoso discernimento sobre certos aspectos do movimento e da locomoção de organismos extintos. As oportunidades foram largamente expandidas com a facilidade de escaneamento e composição de imagens de estruturas 3-D, como ossos e conchas. Essas imagens podem então ser testadas utilizando softwares padrão de engenharia para determinar como a estrutura geral do esqueleto era modelada pelo estresse e a tensão do caminhar, correr, alimentar ou ‘bater cabeças’. Uma abordagem útil de modelagem é a “finite element analysys”ou FEA, um método bem estabelecido e utilizado por engenheiros para acessar a força de pontes ou outras edificações antes de sua construção. Agora ela é também aplicada para entender a estruturação de crânios de dinossauros, entre outras coisas.
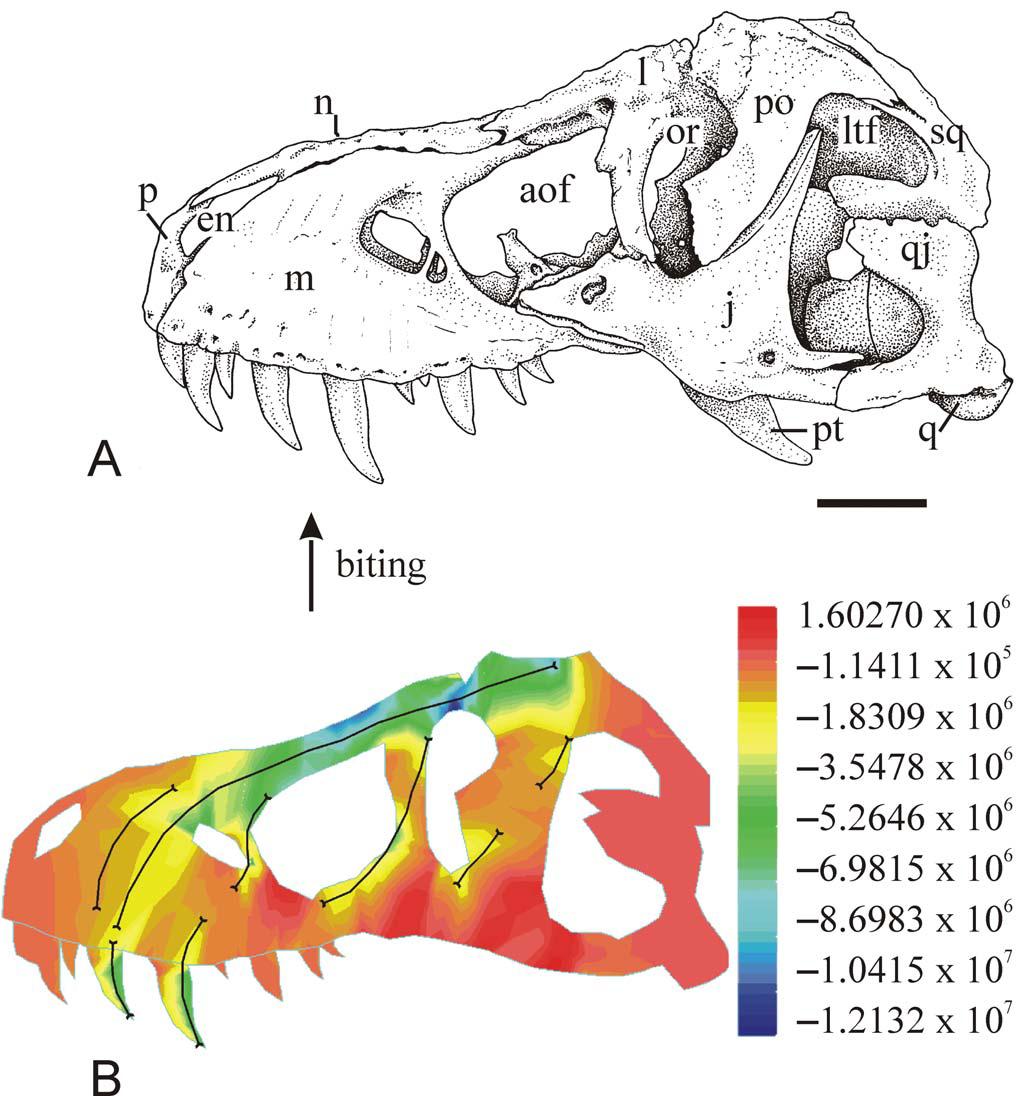
“Finite element analisys“, ou FEA, do crâneo de T. rex
Uma série de tentativas foram feitas para entender como os dinossauros corriam ou caminhavam, e claro, muitas delas foram focadas no T. rex. É razoável assumir que as leis da física e os princípios da biomecânica eram os mesmos no passado, assim como são hoje. Por exemplo, o ponto de partida de um estudo de locomoção para qualquer animal, especialmente um bípede, é estabelecer o seu centro de massa – aconteça o que acontecer o animal não pode cair à toa. O centro de massa para um animal vivente ou extinto pode ser determinado tanto por meio de modelos sólidos como por cálculos de distribuição de tecidos e espaços vazios utilizando-se as fatias do corpo reconstruído em 3-D. Em T. rex, o centro de massa está logo à frente dos quadris e a cauda balanceia o corpo, dando a postura mais natural ao animal com sua coluna posicionada quase totalmente na horizontal. Isso é um maravilhoso progresso sobre a maneira ‘canguru’ como os dinossauros eram representados, com o corpo quase totalmente na vertical e a cauda arrastando no chão. Porém, isso é só o começo, baseado nos mesmos fundamentos T. rex pode ser reconstruído assumindo diversas poses ao se caminhar ou correr.
Ok, e qual seria a velocidade de T. rex? Várias estimativas já foram feitas. Algumas calcularam cerca 20 metros por segundo (72 km/h), enquanto outras, algo em torno de 5 metros por segundo (18 km/h) – a velocidade de um corredor de longa-distância humano. Muitos tipos diferentes de aproximações foram utilizadas para se chegar a isso, e várias ilustram a ingenuidade de alguns paleobiólogos. Por exemplo, pistas de pegadas podem ajudar a indicar a velocidade, já que existe uma relação constante entre o espaçamento das pegadas, o comprimento da perna do animal e a velocidade desenvolvida. Mas existem ainda outras formas de aproximação: as baseadas no comprimento relativo dos ossos das pernas, ou na suposição do risco de injúria para o animal, caso ele caia, utilizando-se de cálculos de estresse e tensão (quanto mais rápido o animal correr, maior o impacto do pé sobre o solo). Recentemente, Ponzer et al. (2009), finalmente sintetizaram toda a história. Eles utilizaram cálculos baseados na estimativa do volume muscular das pernas do animal (os maiores músculos das pernas que alimentam as passadas são proporcionais à massa total e a velocidade do animal). Ponzer e colaboradores concluíram que em velocidade de 5 metros por segundo, o T. rex de 6 toneladas necessitaria de músculos proporcionais àqueles de uma galinha, já se atingisse velocidade de 20 metros por segundo, a maior já assumida para o animal, os músculos da perna do T. rex deveriam corresponder a mais de 86% de sua massa corpórea total. Recalculando isso de maneira a considerar a biomecânica da corrida, as taxas metabólicas envolvidas e evidências de tetrápodes atuais, foi possível concluir que dinossauros de grande porte deveriam exceder as capacidades máximas aeróbicas dos ectotermos modernos. Isso significa que eles seriam funcionalmente endotermos, mesmo que essa ‘homeotermia’ tenha origem por inércia térmica devido ao grande tamanho desses animais.
Frente a tudo o que foi discutido aqui, já é o suficiente para concluir que não há a necessidade de que os paleobiólogos façam especulações desvairadas para reconstituir o comportamento e a função de organismos do passado. Existem metodologias bem fundamentadas para se chegar às hipóteses. E as hipóteses estão sim sujeitas à refutação em qualquer momento. Interpretações engenhosas e descobertas espetaculares, como a de Sanajeh no ninho de saurópodes, podem diversas vezes nos fornecer extraordinários insights sobre a vida perdida do passado e isso nos ajuda a compreender como cada organismo evoluiu (não só anatomicamente) para se tornar o que é hoje.
Benton, M. J., 2010. Studying function and behavior in the fossil record. PLos Biol, 8(3): e1000321:doi10.1371/journal.pbio.1000321
Pontzer, H., Allen, V. & Hutchinson, J., 2009. Biomechanics of running indicates endothermy in bipedal dinosaurs. PLos One, 4(11): e7783.doi:10.1371/journal.pone.0007783.
Zhang, F.; Keams, S.; Orr, P.; Benton, M.; Zhou, Z. et al., 2010. Fossilized melanosomes and the color of Cretaceous dinosaurs and birds. Nature. E-pub ahead of print. doi:nature08740.3d.
Wilson J.; Mohabey, D., Peters, S & Head, J., 2010. Predation upon hatchling dinosaurs by a snake from Late Cretaceous of India. PLoS Biology, 8(3): e1000322.doi:10.1371/journal.pbio.1000322.
Witmer, L., 1995. The extant Phylogenetic Bracket and the importance of reconstructing soft tissue in fossils. In: Thomason, J. ed. Funcional Morphology in Vertebrate Paleonology. New York: Cambridge University Press. pp 19-33.



