

Por Matheus P. dos Santos da Rocha & Cledston Matheus A. Macário
Quando falamos em Paleontologia, muitos a resumem como uma ciência meramente de descrição de aspectos morfológicos, como o simples trabalho de encontrar um osso, descreve-lo e, por sorte, dar nome a uma nova espécie. Porém, a Paleontologia vai muito além disso. Por meio dela, podemos especular sobre diversos aspectos da vida no passado. Até mesmo alguns cujas evidências, muitas vezes, são escassas no registro fossilífero. Um exemplo disso, seria encontrar uma resposta para a pergunta: como eram os olhos dos dinossauros não-avianos?
Como toda ciência, a paleontologia trabalha, inicialmente, com hipóteses, e essas, podem nos levar para linhas de raciocínio beeeem inusitadas, uma hora podemos estar debatendo sobre buracos negros e a extinção dos dinossauros e isso, mais à frente, pode terminar numa deliciosa (ou não) receita de macarrão com biscoito. A história de hoje começa com uma linhas de raciocínio inusitadas: ela parte de um grupo de insetos fósseis, os Kalligrammatidae…
O que são os Kalligrammatidae?

Você já viu em algum jardim por aí pequenas bolinhas sustentadas por um fio bem fino, presas nas folhas das plantas? Se sim, com quase toda certeza você viu ovos de bicho-lixeiro. Pertencentes a uma ordem de insetos chamada Neuroptera, esses inofensivos (para os humanos) insetos são predadores vorazes de ovos de aranhas e outros invertebrados. Essa ordem inclui desde a formiga-leão, até coisas estranhas como os mantispídeos (que parecem uma mistura bizarra entre um marimbondo e um louva-a-deus).
Apesar de não serem um grupo muito comum nos dias de hoje, a representação fóssil deles é abundante. No Brasil, dados de um trabalho de revisão de 2018, dão conta que das 379 espécies de insetos descritos para a Formação Crato, da Bacia do Araripe, 76 são neurópteros, ou seja, 20% da diversidade de insetos da formação está em uma única ordem, que atualmente representa 0,6% das espécies de insetos viventes.
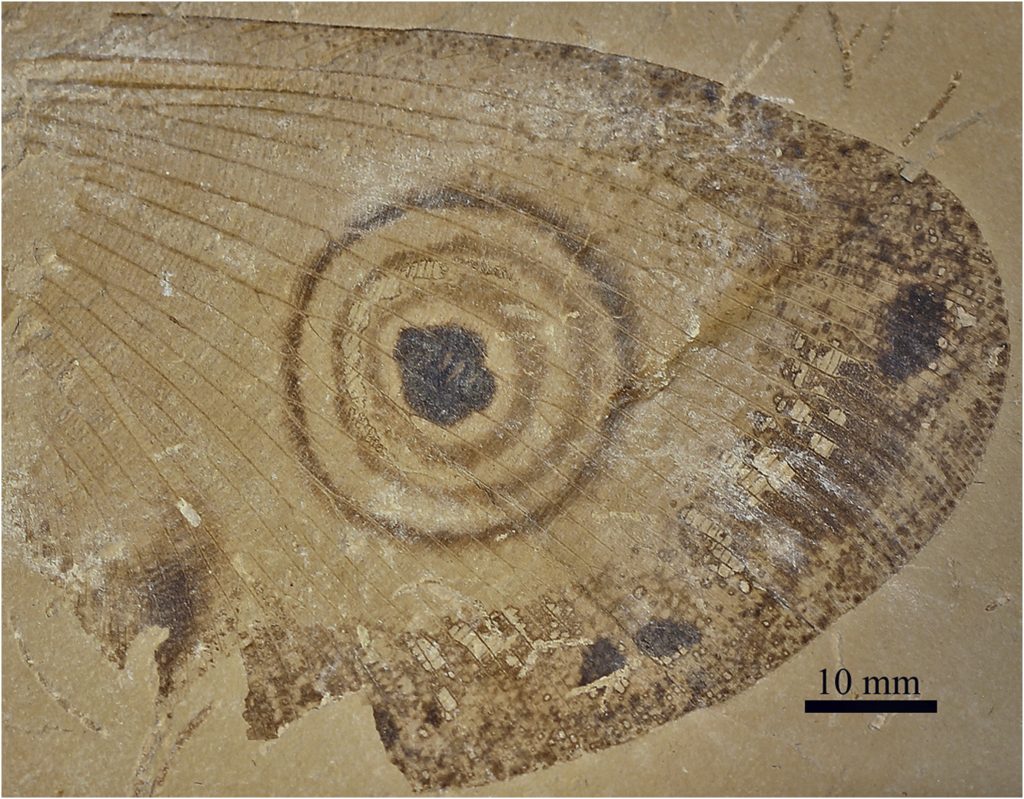
No meio de toda essa diversidade, os fósseis mais enigmáticos de Neuroptera são os da família Kalligrammatidae. O primeiro de Kalligrammatidae foi descrito por Johannes Walther, em 1904, com base num material quase completo, encontrado no calcário jurássico de Solnhofen (Alemanha) – aquele mesmo do Archaeopteryx. Desde então, diversas espécies de Kalligrammatidae foram encontradas em várias localidades, com destaque para os achados na China e nos âmbares birmaneses, ao norte de Mianmar.

Em 1997, o lendário paleontólogo Rafael G. Martins-Neto, descreveu, pela primeira vez, um Kalligrammatidae na Formação Crato, batizado de Makarkinia adamsi. De lá pra cá, outros trabalhos confirmaram a presença dessa família no Nordeste Brasileiro e, inclusive, descreveram novas espécies, sendo este, até hoje, o único lugar fora da Europa e Ásia a ter esses registros.
É uma sorte que esses animais ocorram em vários afloramentos do tipo lagerstätten (sítios com preservação excepcional) pelo mundo afora. A boa preservação dos fósseis permitiu notar rapidamente a semelhança dos kalligrammatídeos fósseis com as atuais borboletas e mariposas. Essa comparação não fica só por conta do formato, padrões de coloração e desenhos das asas, mas alguns espécimes bem preservados, principalmente em âmbar, mostram também a presença de uma “boca” modificada em um fino e comprido tubo chamado de probóscide, característica marcante das mariposas e borboletas (ambas pertencentes à ordem Lepidoptera). Mas isso aconteceu nos kalligrammatídeos num momento do tempo geológico em que as borboletas não existiam e as mariposas não eram tão abundantes e diversificadas como são hoje.
Fora do Brasil, alguns Kalligrammatidae chegam a ser apelidados de “giant lacewings” (crisopídeos gigantes) e isso chegou ao extremo em algumas espécies fósseis. Comparativamente, algumas espécies fósseis são enormes em relação aos seus irmãos ainda viventes. Estima-se que as espécies encontradas no Araripe, por exemplo, alcançavam entre 24 a 32 centímetros de envergadura!
A história dos “olhos” nas asas
Insetos grandes e chamativos podem virar comida facilmente, por isso, precisam ter alguma forma de se proteger da predação. Os kalligrammatídeos que viveram entre o Eojurássico ao Neocretáceo estavam dividindo espaço com lagartos, dinossauros avianos e não-avianos, pterossauros, entre outros predadores . Logo, teria que haver alguma forma deles não sucumbirem a seus colegas de habitat!
As mariposas e borboletas de hoje em dia têm algumas estratégias para evitar a predação. Desde projeções nas asas para desviar a atenção do predador, como as mariposas do gênero Actias, até mimetizar (imitar) folhas secas, tal qual Zaretis itys faz. Outra forma é ter “olhos”, ou melhor, ocelos em suas asas. Os ocelos são desenhos circulares que aparecem em diversos animais, especialmente nos lepidópteros. Esses círculos podem aparecer com 2 estratégias diferentes de uso:

- A primeira é ter eles próximos às margens da asa, fazendo com que a atenção de um provável predador seja focada na ponta da asa e não no centro do corpo do organismo.
- A outra é simplesmente aterrorizar! As mariposas da família Saturniidae e as borboletas-olho-de-coruja do gênero Caligo, por exemplo, fazem isso muito bem. Elas têm ocelos enormes no centro das asas, que imitam – algumas vezes de forma assustadora – os olhos de uma coruja, afastando assim qualquer predador que ouse atacá-las.

E é nesse ponto que queríamos chegar. Justamente essa segunda estratégia é atribuída a várias espécies fósseis de kalligramatídeos. Desde o primeiro espécime descrito, os ocelos gigantes estão presentes nas asas, e há trabalhos que descrevem e comparam os diversos formatos encontrados.
O que isso tem a ver com dinossauros?
Agora, chegou a hora que, ou vocês sairão desse blog nos chamando de loucos, ou terão o famoso “Mind Blow”. Vamos ao ponto principal: você já parou para pensar sobre o formato dos olhos dos dinossauros não-avianos? Essa é uma discussão complicada, pois o número de olhos de dinossauro preservados no registro fossilífero é: zero! Mas é uma curiosidade legítima querer saber essa informação, tanto que pode ser encontrado por aí, em fóruns pela internet, pessoas debatendo sobre essa questão.
Como esse tipo de material fóssil para dinossauros é inexistente, parte-se para a comparação com animais recentes, tanto seus parentes mais próximos ainda vivos, quanto possíveis análogos ecológicos. Mas existe ainda outra linha de raciocínio para se debater: não olhar para os dinossauros em si, mas para seus colegas de habitat e, no nosso caso especifico, os kalligramatídeos da Formação Crato.

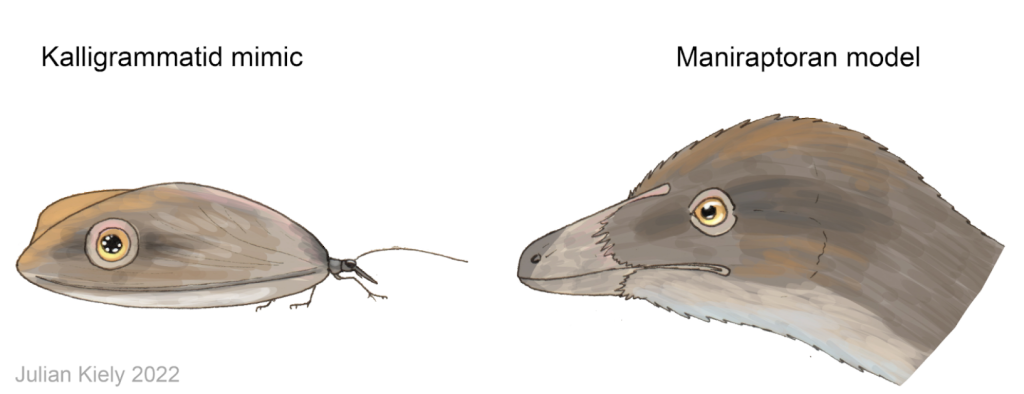
Como já foi mencionado anteriormente, os “Giant Lacewings” poderiam ter se utilizado da segunda estratégia de uso dos ocelos: para assustar prováveis predadores, imitando os olhos de animais com os quais conviveram. Aí está o “pulo do gato”. Para um predador se assustar com os olhos desenhados nas asas das borboletas-olho-de-coruja é preciso que tenha um animal no mesmo habitat, que vá servir de gatilho (o “modelo” dos ocelos de Caligo, uma coruja, por exemplo: um predador assustador, que assuste o predador da Caligo). Mas há 120 milhões de anos não existiam corujas no Ceará, então…quem eram os modelos dos Kalligrammatidae do Crato?

Existem dois principais suspeitos: pterossauros e dinossauros, mas vamos por partes. Pterossauros na Bacia do Araripe, segundo Mendes et al. (2020), eram majoritariamente piscívoros (comedores de peixes), com algumas exceções como Lacusovagus magnificens, que provavelmente vagava pelos pântanos da região para caçar anfíbios e outras pequenas presas. O trabalho de Mendes, inclusive, coloca os pterossauros como animais no topo da teia trófica da região na época.
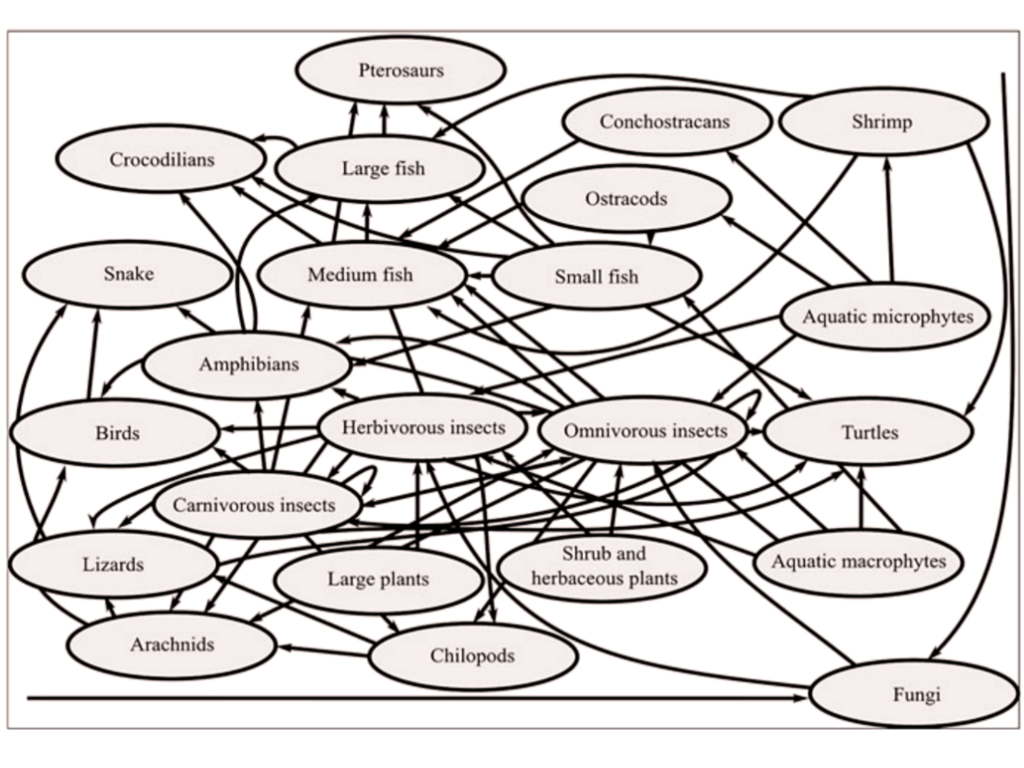
Mas se os pterossauros cearenses comiam peixes, majoritariamente, os possíveis predadores dos kalligramatídeos (outros insetos, anfíbios, pássaros, pequenos dinossauros, etc.) não estavam no cardápio deles, a priori. Por esse fator, seria compreensível a exclusão desses animais como possíveis modelos para os ocelos.

Já os dinossauros, por outro lado, são os candidatos perfeitos para esse quebra-cabeças ecológico. Animais como Aratasaurus museunacionali, Mirischia asymetrica (que, assim como “Ubirajara“, foi traficado para Alemanha #MirischiaBelongtoBR) e Santanaraptor placidus, ocupavam o nicho de predadores de médio a pequeno porte da região do Cariri. Como apontado por Julian Kiely, em seu artigo para o blog “Paleoflora”, a forma da asa dianteira na maioria das espécies de kalligramatídeos, e o grande tamanho dessas asas, correspondiam, aproximadamente, ao tamanho e a forma das cabeças de muitos pequenos dinossauros predadores que conviviam com esses insetos (como as espécies mencionadas acima). Desta forma, poderíamos inferir que as pupilas dos dinossauros de médio a pequeno porte do Jurássico Superior e do Cretáceo Inferior, como os maniraptores (pelo menos), deveriam ser arredondadas, já que os ocelos de todos os kalligramatídeos conhecidos até então, possuem esse mesmo formato. O que se soma à evidência indireta parelela, que considera como base comparativa o formato da pupila dos dinossauros viventes, que são as aves.
O poder da especulação
Alguns podem estar se perguntando: qual a importância de especular aspectos biológicos e evolutivos tão difíceis de se comprovar por meio do registro fossilífero? Muito da ciência começa com especulação. As descobertas científicas, em geral, nascem de hipótese de alguém. Um exemplo clássico foi a detecção das ondas gravitacionais em 2015, que haviam sido previstas por Albert Einstein em 1916.
Focando na área da Paleontologia, um exemplo muito interessante, e, à época, considerado extremamente especulativo, foi o da existência de um radiodonte (grupo que inclui o Anomalocaris) filtrador, batizado de “Ceticaris”. Nada mais que uma especulação concebida pelo artista John Meszaros, publicada no livro All Your Yesterdays, de 2013. Porém, para surpresa de muitos, em 2014 foi realmente descrito um radiodonte cambriano com hábito filtrador, Tamisiocaris borealis.

Em homenagem à previsão de Meszaros, Tamisiocaris foi incluído em um novo clado denominado Cetiocaridae. Infelizmente, o nome deste clado não é mais considerado válido, de acordo com o Código Internacional de Nomenclatura Zoológica, por não existir nenhum gênero real chamado “Cetiocaris“, então foi formalmente substituído pelo nome Tamisiocarididae.

Finalmentes
Com base em todos os argumentos supracitados (Alô, professores de redação!), podemos inferir que a hipótese levantada pode levar a especulações e trabalhos futuros que respondam às nossas dúvidas (isso claro, se esse post, neste humilde blog, chegar nas pessoas certas, e para isso seu compartilhamento é fundamental). Gostaríamos de agradecer a Julian Kiely do excelente blog “Paleoflora” pelo artigo que inspirou este, e que isso inspire a todos os nossos leitores a imaginar e especular dentro da ciência, lançar ideias, compartilhá-las, pois só assim a ciência cresce e prospera, com união e partilha.
Referências:
Martins-Neto, R. G. 1997. Neurópteros (Insecta, Planipennia) da Formação Santana (Cretáceo IInferior) Bacia do Araripe, Nordeste do Brasil. X – descrição de novos taxa (Chrysopidae, Babinskaiidae, Myrmeleontidae, Ascalaphidae e Psychopsidae). Revista Universidade Guarulhos , São Paulo, v. 2, n.4,. p. 68-83.
Kiely, J. 2022. Restoring the Kalligrammatids: The not-butterflies of the mesozoic. Paleoflora.





















{kind=link}