

Texto por Thiago Carlisbino
Bem no início da história evolutiva dos tetrápodes, inúmeros animais eram adaptados para a sobrevivência no ambiente aquático. Os ditos anfíbios, na verdade, compunham uma grande diversidade de animais (ex. temnospôndilos, lepospôndilos – Fig. 1), que aterrorizavam áreas inundadas e outros corpos d’água. No entanto, nesse ambiente hostil, surgiram pequenos e frágeis tetrápodes com uma especialização reprodutiva inovadora: o ovo amniótico.
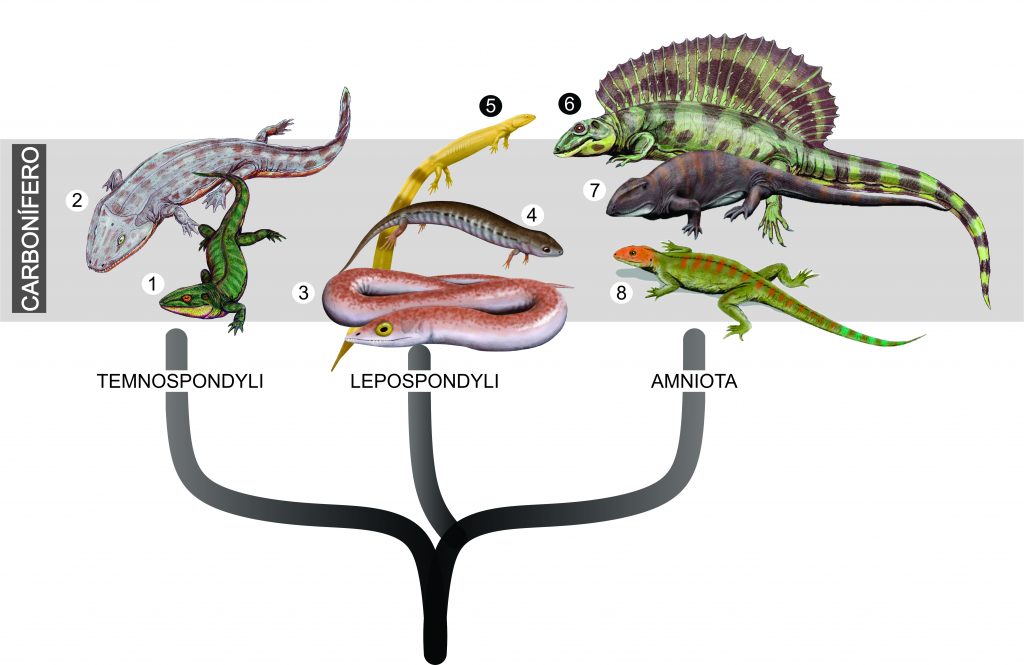
Geralmente é ensinado que a presença de membranas diferenciadas e externas ao embrião fizeram com que os tetrápodes conquistassem definitivamente o ambiente terrestre e se diversificassem em formas gigantescas e até mesmo voadoras. Dessa forma, os Amniota ganharam o título de colonizadores do ambiente terrestre. Mas, e o ambiente aquático? Ficou para os anfíbios? Os amniotas deram as costas para o ambiente aquático?
A resposta é NÃO e no Brasil temos um dos grupos fósseis mais importantes para contar essa história: os mesossauros!
Mesossauros?
Talvez os Mesosauridae sejam os amniotas melhor representados e conhecidos no registro fossilífero do nosso país. Seus fósseis são abundantes em rochas permianas da Formação Irati e sua ampla distribuição geográfica (de Goiás até o Rio Grande do Sul) garantiu um bom conhecimento acerca do seu modo de vida e aspectos osteológicos, principalmente da região pós-craniana (Modesto, 2009; Sedor & Ferigolo, 2001).
Os mesossauros fazem parte de um grupo de amniotas conhecido como Parareptilia. Os “pararrépteis” formam uma pequena linhagem que surgiu no Carbonífero e apresenta uma grande diversidade de formas com especializações variadas e novidades evolutivas. Para vocês terem ideia da importância do grupo, nos pararrépteis encontramos: formas herbívoras de grande porte (como os pareiassauros); o mais antigo réptil com capacidade de locomoção bípede conhecido (Eudibamus cursoris); os primeiros amniotas com implantação dentária do tipo tecodonte (dentes implantados em alvéolos, e.g. Bolosaurus grandis) e o primeiro grupo de amniotas a apresentar especializações para o ambiente aquático: sim, os mesossauros.
As características que indicam que esses animais eram adaptados para a vida no meio aquático são bastante evidentes (Fig. 2): eles apresentam crânio com rostro alongado (i.e. focinho comprido), narinas próximas às órbitas, coluna vertebral longa, principalmente na região caudal, pés espalmados e ossos com paquiostose e osteosclerose (explico a seguir).
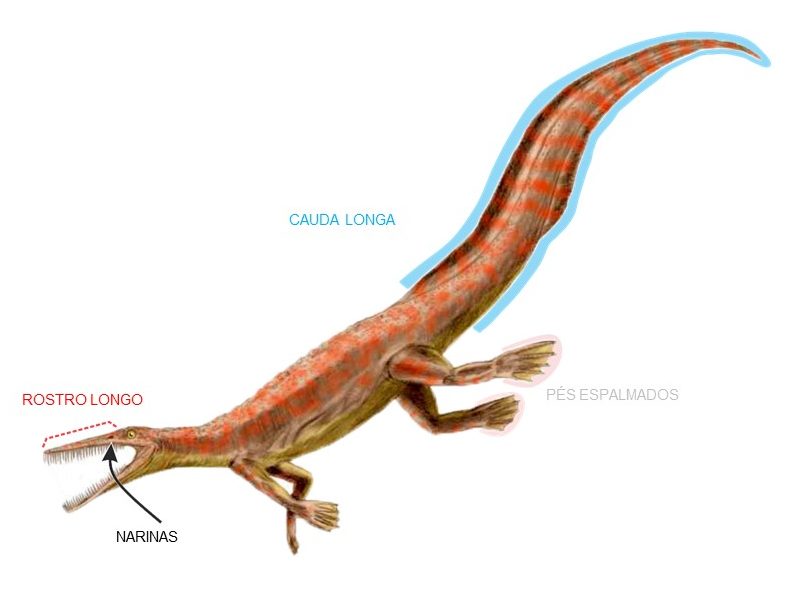
Figura 2. Representação artística de Mesosaurus tenuidens. Ilustração por Nobu Tamura (Creative Commons).
Os mesossauros viviam em um mar extenso (mar Irati-Whitehill) confinado, que cobria áreas que atualmente correspondem ao Brasil, África do Sul, Paraguai e Uruguai (Oelofsen & Araújo, 1983). Nesse grande corpo d’água, os mesossauros compartilhavam o ambiente com… bom, com nenhum outro tetrápode! É isso mesmo! Até o momento, esses pararrépteis são os únicos tetrápodes conhecidos para a Formação Irati. Curioso né?
Na unidade, também ocorrem outros vertebrados (e.g. Chahud & Petri, 2009; 2010), tais como fósseis de peixes (tubarões, celacantos e paleonisciformes) e um dente de um provável anfíbio, mas em camadas de rocha distintas, tornando muito difícil inferir que esses vertebrados compartilhavam o mesmo ambiente numa mesma época.
Se eles não compartilhavam o ambiente com outros vertebrados, então os dentes compridos dos mesossauors eram usados para que?
Entre os mesossaurídeos são reconhecidas três espécies (Fig. 3): Brazilosaurus sanpauloensis, Stereosternum tumidum e Mesosaurus tenuidens. Nem todos os mesossauros apresentavam dentes compridos. B. sanpauloensis, por exemplo, apresenta os dentes mais curtos entre os Mesosauridae, mostrando que haveria uma certa variação de dieta entre eles. Sobre a alimentação dos mesossauros, temos fortes pistas: fósseis de crustáceos são encontrados em abundância nas mesmas rochas que nossos pararépteis. Dessa forma, é inferido que pelo menos M. tenuidens se alimentasse desses crustáceos, sendo os dentes utilizados como uma espécie de filtro, que prenderia os invertebrados. Além disso, alguns pesquisadores também sugerem canibalismo e necrofagia, devido a presença de pequenos ossos encontrados em conteúdos gástricos e coprólitos associados de mesossaurídeos (Silva et al. 2017).

Voltando ao assunto dos ossos: Paquiostose? Osteosclerose? O que é isso?
O osso é um tecido vascularizado, composto por células e uma matriz extracelular mineralizada. É um tecido vivo, que tem a função de suporte, proteção, produção de células sanguíneas e também atua como regulador do equilíbrio mineral do organismo (Hall, 2005).
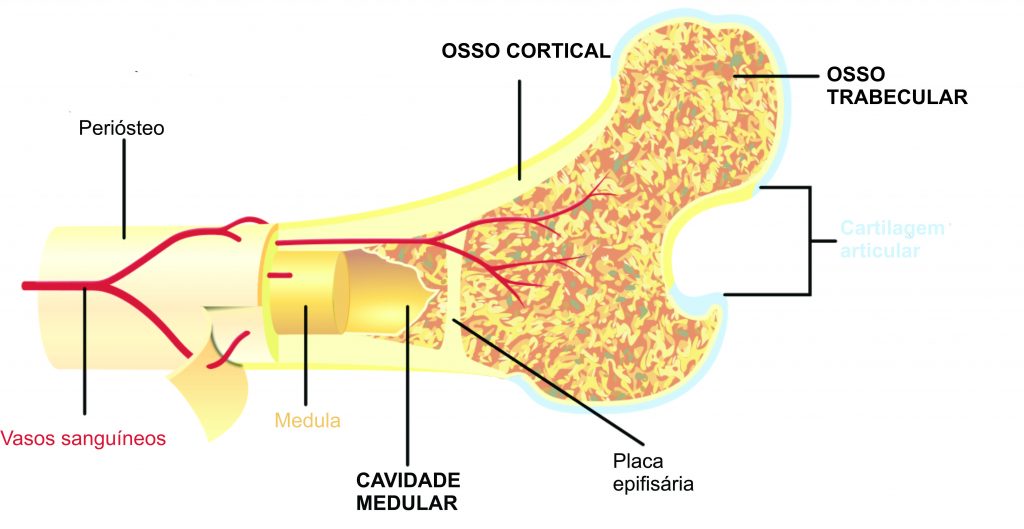
Se pegarmos um osso longo (ex.: fêmur, úmero ou costela) e fizermos um corte na porção média dele, é bem provável veremos uma parte externa mais compacta e outra interna menos mineralizada, mais porosa (variações existem, é claro). A porção periférica é chamada de córtex (osso cortical na imagem), enquanto a interna é chamada de cavidade medular (que contém a medula óssea) (Fig. 4). Em estudos microanatômicos, são detalhadas as variações dessas regiões nos ossos, tais como a espessura do córtex, diâmetro do canal medular e porosidade. O aspecto microanatômico do osso é importante para entender a paleobiologia do vertebrado fóssil, pois sua organização está intimamente relacionada com o hábitat, modo de vida e aspectos biomecânicos do organismo (Krillot et al. 2008). Portanto, é uma ferramenta útil para quem deseja entender como determinado animal vivia em um passado distante.
Para exemplificar, vou tentar caracterizar os aspectos microanatômicos de ossos longos em espécies terrestres:
Nestas formas, a relação entre espessura cortical, diâmetro do canal medular e seu preenchimento ou não por trabéculas ósseas, parece depender muito do porte do animal.
No geral, amniotas terrestres apresentam ossos dos membros com córtex delgado, canal medular amplo e sem a presença de trabéculas. No entanto, a situação muda quando o animal se torna muito grande. Imaginem um osso extremamente comprido e com poucos milímetros de espessura cortical: será que ele aguentaria suportar toneladas e ao mesmo tempo resistir ao estresse mecânico provocado pela locomoção? É bem provável que não. Dessa forma, é comum que o osso desses animais apresente córtex espesso e cavidade medular preenchida por osso trabecular (i.e. em forma de travas) (Fig. 5).
Já a vida no meio aquático impõe uma série de restrições mecânicas diferenciadas, devido à água apresentar maior densidade e viscosidade que o ar. Nos ossos é comum o aumento em massa e densidade, que ocorre devido à grande quantidade de deposição de osso cortical e o aumento da compacidade interna. Dessa forma, ossos paquiostóticos possuem aspecto inchado, devido à grande quantidade de osso cortical depositado, enquanto que ossos osteoscleróticos são extremamente compactos, quase não apresentando canais vasculares e, quando apresentam, são geralmente de pequeno calibre. Nos mesossaurídeos, tanto a paquiostose, quanto a osteosclerose ocorrem em vértebras e costelas (Fig. 6). Tal característica é denominada de paquiosteosclerose.
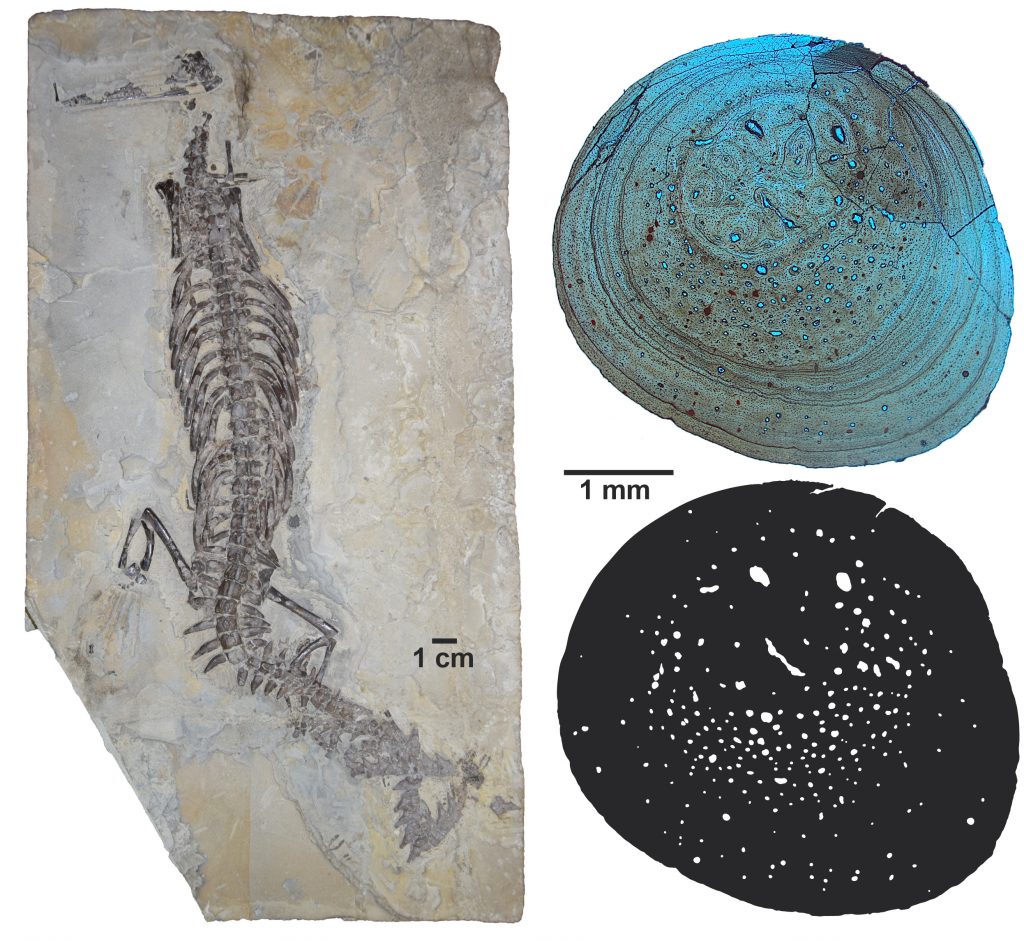
No entanto, essa adaptação tem um preço: o aumento da massa esquelética tem consequências na locomoção subaquática (Taylor, 2000). A paquiostose parece ocorrer com mais frequência em espécies que habitam áreas rasas aonde o animal tem a possibilidade de alcançar o fundo com certa facilidade. Táxons de uma mesma linhagem, mas adaptados ao ambiente pelágico (que vivem em mar aberto) tendem a não apresentar tal característica o que não acontece em formas “menos ágeis” (Houssaye, 2009). De fato, os mesossauros não alcançavam grande velocidade na água e, além disso, os indivíduos encontrados em rochas depositadas em ambiente mais profundo (ex. folhelhos), são geralmente os de maior tamanho (Oelofsen & Araújo, 1983; Villamil et al. 2015).
Além dos aspectos ósseos, há outras evidências que sugerem o habitat preferencial dos mesossauros?
Além dos fósseis de mesossauros ocorrerem apenas em rochas que foram depositadas em ambiente marinho, também temos evidências icnológicas. Sedor & Silva (2004), descreveram pela primeira vez marcas subaquáticas de mesossaurídeos em calcários da Formação Irati no Estado de Goiás. As marcas foram produzidas pelo toque dos pés no sedimento durante o impulso para aumento de velocidade ou mudança de curso. Posteriormente, Silva et al. (2009) estudaram pegadas de mesossauros semelhantes de material oriundo do Estado do Paraná e descreveram a icnoespécie Mesosaurichnium natans (Fig. 7).

Apesar do estudo microscópico dos fósseis se dar a partir de métodos destrutivos (lembrem-se de ter muito cuidado na escolha dos espécimes!), ele fornece detalhes muito interessantes acerca da paleobiologia e paleoecologia dos táxons fósseis. Neste texto, descrevi brevemente o que podemos inferir a partir dos detalhes microanatômicos dos ossos. No entanto, existem outros aspectos que podemos interpretar a partir de lâminas delgadas de fósseis: detalhes do crescimento, a taxa de deposição óssea e a possibilidade de se inferir a idade de um indivíduo são apenas algumas delas.
Nos mesossauros a análise da microestrutura óssea ainda está no início e tem muita coisa para ser mostrada.
Até mais!
—————————
Quer saber mais sobre esses fofos? Corre lá no canal pra conferir dois vídeos super bacana que a gente fez sobre eles!
Referências
CHAHUD, A., PETRI, S. (2009). Novos Xenacanthidae (Chondrichthyes, Elasmobranchii) da base do Membro Taquaral, Formação Irati, Permiano da Bacia do Paraná. Revista do Instituto Geológico, 30(1-2), 19-24.
CHAHUD, A., PETRI, S. (2010). Anfíbio e Paleonisciformes da Porção Basal do Membro Taquaral, Formação Irati (Permiano), Estado de São Paulo, Brasil. Geologia USP: Série Científica, 10(1), 29-37.
HALL, B. K. (2005). Bones and cartilage: Developmental and and evolutionary skeletal biology. Elsevier, 2005, 792p.
HOUSSAYE, A. (2009). “Pachyostosis” in aquatic amniotes: a review. Integrative Zoology, 4 (4), 325-340.
HOUSSAYE, A. et al. (2015). Biomechanical evolution of solid bones in large animals: a microanatomical investigation. Biological Journal of the Linnean Society, 117, 50-371.
KRILLOF, A. et al. (2008). Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference. Journal of Evolutionary Biology, 21, 807-826.
MODESTO, S. P. (2010). The postcranial skeleton of the aquatic parareptile Mesosaurus tenuidens from the gondwanan Permian. Journal of Vertebrate Paleontology, 30(5), 1378-1395.
OELOFSEN, B. W. & ARAÚJO, D. C. (1983). Palaeoecological implications of the distribution of mesosaurid reptiles in the Permian Irati Sea (Paraná Basin), South America. Revista Brasileira de Geociências, 13, 1-6.
SEDOR, F. A. & FERIGOLO, J. (2001). A coluna vertebral de Brazilosaurus sanpauloensis Shikama & Ozaki, 1966 da Formação Irati, Permiano da Bacia do Paraná (Brasil) (Proganosauria, Mesosauridae). Acta Biologica Paranaense, 30, 151-173.
SEDOR, F. A. & SILVA, R. C. (2004). Primeiro registro de pegadas de Mesosauridae (Amniota, Sauropsida) na Formação Irati (Permiano Superior da Bacia do Paraná) do Estado de Goiás, Brasil. Rev. Bras. Paleontol. 7, 269–274. doi: 10.4072/rbp.2004.2.21
SILVA, R. C., SEDOR, F. A., FERNANDES, A. C. S. (2009). Ichnotaxonomy, functional morphology and paleoenvironmental context of Mesosauridae tracks from Permian of Brazil. Rev. Bras. Geociências. 39, 705–716.
SILVA, R. C. & SEDOR, F. A. (2017). Mesosaurid swim traces. Frontiers in Ecology and Evolution, 5: 22.
SILVA, R. R. et al. (2017). The feeding habits of Mesosauridae. Frontiers in Earth Science, 5: 23.
TAYLOR, M. A. (2000). Functional significance of bone ballast in the evolution of buoyancy control strategies by aquatic tetrapods. Historical Biology 14, 15–31.
VILLAMIL, J. et al. (2015). Optimal swimming speed estimates in the Early Permian mesosaurid Mesosaurus tenuidens (Gervais, 1865) from Uruguay. Historical Biology, 28, 963-971.
