

Anteriormente falamos aqui sobre um incrível golfinho caçador que dominou as águas do Oligoceno no sul da Califórnia (E.U.A) e suas contribuições para a evolução dos cetáceos (grupo que inclui as baleias e golfinhos). Citamos algumas diferenças entre ele e um grupo de cetáceos muito mais antigo, que incluía também caçadores tão excepcionais quanto, os gigantescos basilossaurídeos. Mas afinal, quem são esses cetáceos com nome de dinossauro?

Os fósseis mais antigos de basilossaurídeos são encontrados em rochas datadas do final do Eoceno Médio. Eles são encontrados em todo o mundo, incluindo África, Ásia, Europa, América do Norte e do Sul, Nova Zelândia e até mesmo na Antártica. De maneira geral, basilossaurídeos eram cetáceos com o corpo extremamente alongado, algumas espécies chegando a mais de 18 metros de comprimento. Foi por meio de fósseis desses cetáceos “arcaicos” que paleontólogos tiveram as primeiras pistas de que os golfinhos e baleias se originaram de mamíferos inicialmente terrestres.
O entendimento da ancestralidade terrestre só foi possível graças à preservação de ossos vestigiais das patas traseiras. Estas, eram pequenas e provavelmente semelhantes às nadadeiras peitorais dos golfinhos atuais, ou seja, não se conectavam/articulavam mais diretamente com a coluna. Isso demonstra que basilossaurídeos não conseguiriam mais sustentar o seu próprio corpo em terra e, portanto, seriam os primeiros cetáceos completamente adaptados à vida marinha. Também foram os cetáceos mais antigos a mostrar a migração das narinas da ponta do focinho para o topo da cabeça, como vemos hoje nas baleias e golfinhos modernos.

Grandes répteis?
A primeira espécie de basilossaurídeo foi descrita em 1834 por Richard Harlan, que batizou-a de Basilosaurus, “Lagarto Rei” em grego antigo. Porém, Harlan havia interpretado aqueles fósseis erroneamente como sendo de um grande réptil marinho. Percebendo esse erro, Richard Owen, em 1839, reavaliou os fósseis do Basilosaurus associando-o aos cetáceos, e dando-lhe um novo nome, Zeuglodon, que significa “Dentes de Jugo”, também derivado do grego antigo.
Apesar dessa nova avaliação, o nome original dado por Harlan permaneceu devido às regras internacionais da nomenclatura zoológica, que definem que o primeiro nome dado a uma nova espécie é o que deve ser considerado válido.
Evidências de um grande caçador
Nos últimos anos, descobertas de novos fósseis de basilossaurídeos na região do Egito, no continente africano, revelaram relações nada amigáveis entre algumas espécies do grupo. Alguns fósseis indicam que o gigantesco Basilosaurus isis, um animal que atingia até 18 metros de comprimento, seria predador de uma espécie menor de basilosaurídeo conhecida como Dorudon atrox. Essa descoberta deu pistas sobre como os grandes basilossauros do Eoceno Médio e Final eram efetivamente os “reis” dos mares.

Julia M. Fahlke relata em um trabalho publicado em 2012, que fósseis das duas espécies de basilossaurídeos foram encontrados no mesmo sítio fossilífero. Porém, havia algo estranho ali: na localidade eram encontrados fósseis tanto de juvenis quanto de adultos de Dorudon atrox e somente fósseis de adultos de Basilosaurus isis. No mesmo trabalho, Fahlke relata também, que alguns dos crânios de D. atrox possuíam grandes marcas de mordida, que depois de análise detalhada, puderam ser interpretadas como ferimentos letais, ou seja, elas eram a provável causa de morte desses organismos. Quem seriam os responsáveis pelas mordidas?
Isso levou a pesquisadora a propor a hipótese de que adultos de B. isis poderiam, no passado, ter invadido as áreas de parto de D. atrox para atacar seus filhotes. Não havia, no entanto, nenhuma evidência direta para apoiar essa hipótese. Fahlke, então, decidiu digitalizar os fósseis para testar a sua ideia. Ela aplicou técnicas de tomografia computadorizada e obteve modelos tridimensionais (3D) dos crânios fossilizados. Os modelos 3D dos espécimes juvenis de D. atrox foram colocados digitalmente na boca de um B. isis adulto e as marcas de mordidas comparadas com o tamanho e posicionamento dos dentes. Bingo! As marcas de mordida no crânio dos juvenis de D. atrox correspondiam exatamente à dentição de B. isis.
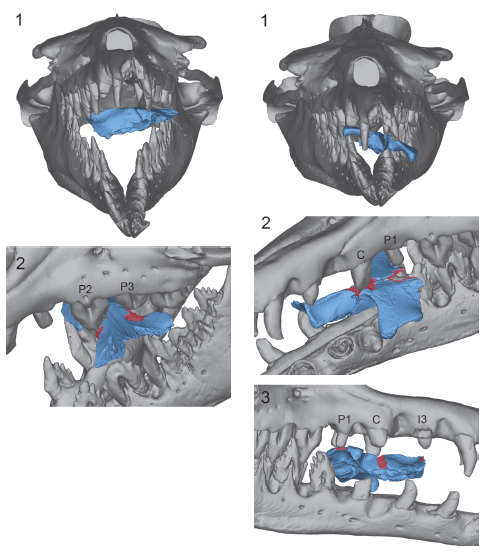
O perfil do culpado
A pesquisadora também produziu modelos de argila dos dentes de B. isis e observou que as marcas nos crânios dos juvenis de D. atrox também se encaixavam com as características específicas dos dentes da espécie maior de basilossaurídeo. As marcas variavam ainda de acordo com qual dente fincou no crânio do animal, a posição do ataque, a força aplicada durante a mordida e o estado de desgaste natural dos dentes de B. isis.
Foi proposto também, que algumas marcas poderiam ter sido causadas por outros animais como Crocodilus megarhinus, grandes crocodilos marinhos do final do Eoceno do Egito, ou Carcharocles sokolowi, um grande tubarão encontrado mesmo depósito, com dentes de até 9,5 cm. Porém, as marcas analisadas não se encaixavam com as características da arcada e dos dentes dessas espécies. O culpado realmente só poderia ser Basilosaurus isis.
Acreditava-se, até então, que a alimentação dos cetáceos primitivos era limitada a peixes, entretanto, com esse estudo, as primeiras evidências de uma predação mais ampla foram sugeridas.
Um mundo onde baleia comia baleia (e também tubarão!)
Apesar do excelente trabalho feito por Fahlke, evidências diretas eram necessárias para comprovar definitivamente a relação de predação proposta pela autora. A preservação do conteúdo estomacal desses enormes predadores seria o ideal.
Eis que, em 2019, Manja Voss e colegas publicaram um trabalho justamente com a peça faltante para a compreensão dessa relação entre presa e predador: o conteúdo estomacal de um B. isis foi encontrado e nele, partes de D. atrox. Voss e colegas relataram evidências de três espécies de vertebrados encontrados no conteúdo estomacal de B. isis. Haviam partes de dois juvenis de D. atrox, dentes de um pequeno peixe ósseo (Pycnodus mokattamensis) e dentes de um grande tubarão (Carcharocles sokolowi).
Os autores sugeriram ainda que, devido ao grande tamanho de algumas presas encontradas, B. isis não deveriam ser capazes de engoli-las inteiras. Considerando o comportamento de cetáceos atuais, eles também não deveriam consumir a carne de carcaças. Eles eram devidamente capazes de atacar animais de grande porte e abatê-los, além de terem dentes apropriados para cortá-los em pedaços. A cena deveria ser terrível.
Muito se comparou B. isis com as grandes orcas (Orcinus orca) da atualidade, pois estas também são de caçadores de topo de cadeia, que consomem animais, incluindo outros mamíferos marinhos e tubarões, às vezes muito maiores do que elas mesmas. A descoberta elucidou um pouco mais sobre a dinâmica dos predadores de topo dos oceanos do início da Era Cenozóica.
As orcas também caçam filhotes de grandes baleias, assim como consomem parte das presas maiores, dando uma perspectiva sobre como B. isis faziam em sua época. Porém, as orcas caçam em grupo e esse tipo de interação não é possível de ser verificada no registro fossilífero de B. isis. Pelo menos até o momento…

Pelas informações adquiridas a partir desses trabalhos, entende-se que B. Isis foram possivelmente os primeiros cetáceos a se alimentarem de outros cetáceos de sua época. Os trabalhos citados aqui também ampliam o conhecimento sobre a evolução da dieta desse grupo de animais, antes interpretados como consumidores exclusivos de peixes. Ainda há muito a se descobrir sobre a paleoecologia dos cetáceos extintos. O registro fossilífero sempre guarda surpresas e é preciso uma atenção especial e, às vezes, até mesmo criatividade para se desvendar os mistérios guardados nos fósseis.
Referências:
FAHLKE, Julia M. Bite marks revisited — evidence for middle-to-late Eocene Basilosaurus isis predation on Dorudon atrox (both Cetacea, Basilosauridae). Palaeontologia Electronica, v. 15, n. 3, p. 32A, 2012.
MARX, Felix G.; LAMBERT, Olivier; UHEN, Mark D. Cetacean paleobiology. John Wiley & Sons, 2016.
VOSS, Manja., ANTAR, Mohammed Sameh., ZALMOUT, Iyad S., & GINGERICH, Philip D. Stomach contents of the archaeocete Basilosaurus isis: Apex predator in oceans of the late Eocene. PloS one, v. 14, n. 1, p. e0209021, 2019.




{kind=link}