Os terópodes são um grupo de dinossauros que surgiu há mais de 200 milhões de anos e habitam todos os continentes ainda hoje. Eles são conhecidos por suas adaptações que os tornaram os mais rápidos e aerodinâmicos de todos os dinossauros. Uma das principais características que contribuíram pra sua agilidade e resistência foi o sistema de sacos aéreos, que permitiu uma respiração mais eficiente e a capacidade de invadir os ossos do esqueleto com câmaras pneumáticas. Esse sistema é fundamental para entender a evolução dos terópodes e sua adaptação pra estes diferentes ambientes.
Hoje vamos falar sobre nosso artigo recente que desvendou um mistério sobre a evolução dos dinossauros terópodes. Este artigo, publicado na revista Journal of Anatomy, investiga a evolução do sistema de sacos aéreos em terópodes, esse grupo de dinossauros que inclui os mais longevos e os únicos que sobreviveram até hoje, na forma das aves.
Recentemente, pesquisadores descobriram que o sistema de sacos aéreos evoluiu pelo menos três vezes de forma independente nos avemetatarsalianos, um grupo que inclui pterossauros, saurópodes e terópodes. Enquanto os saurópodes apresentam uma arquitetura pneumatizada complexa em suas vértebras, os terópodes ainda são menos compreendidos nesse aspecto.
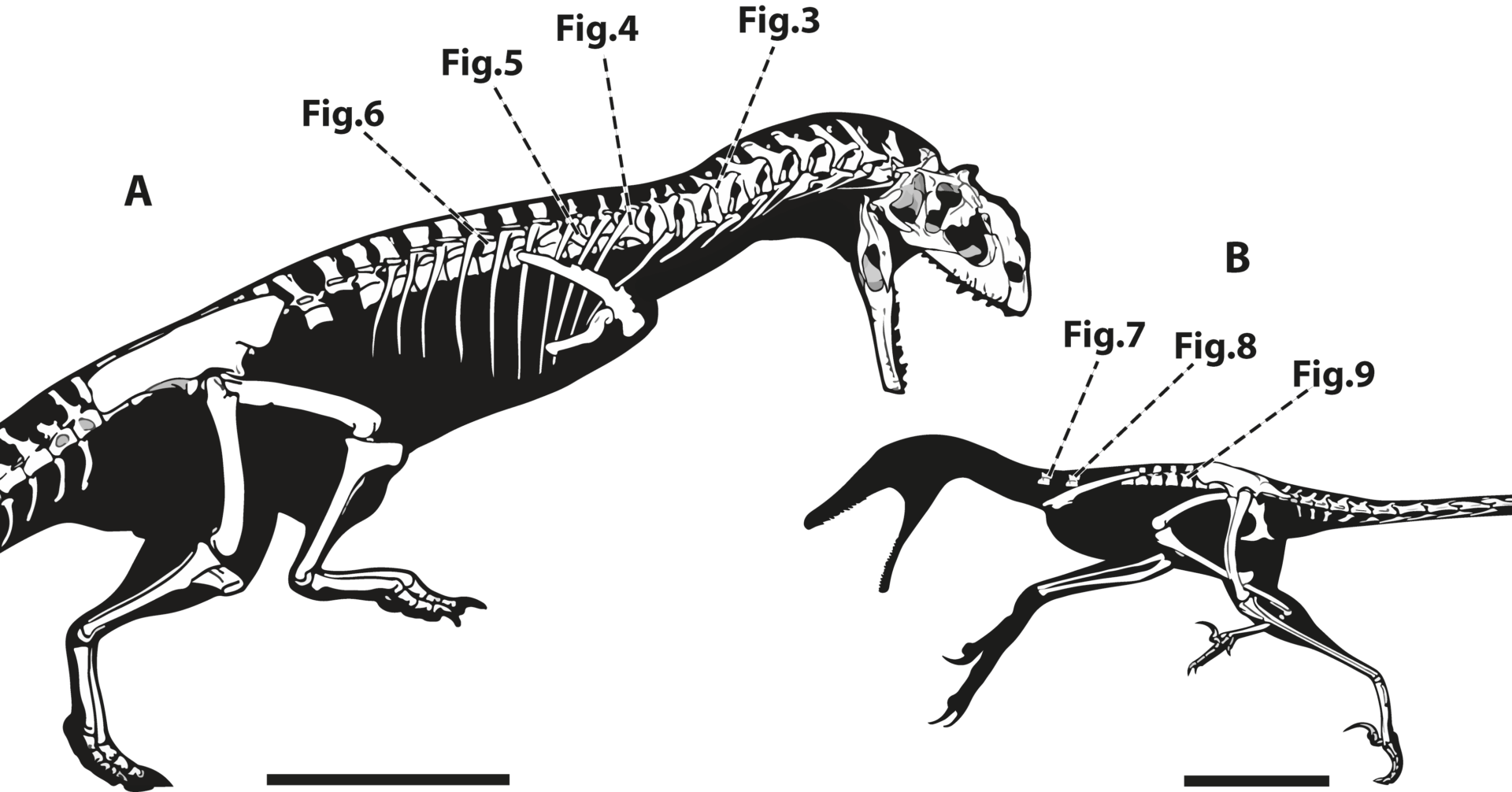
Nesse estudo, nós analisamos o esqueleto axial de dois dinossauros terópodes: Majungasaurus, um ceratosauriano, e Rahonavis, um paraviano. Ambos foram encontrados na Formação Maevarano, no norte da Madagascar, durante expedições realizadas nos últimos 30 anos. Nossa equipe utilizou a tomografia computadorizada desses fósseis pra detectar padrões de pneumatização nos ossos e comparar com outros grupos de terópodes.
Figura apresentando o ceratossauriano Majungasaurus (esquerda) e o paraviano Rahonavis, mostrando os elementos ósseos tomografados. Fonte: Aureliano et al. (2024).
O estudo revelou que o Majungasaurus, um dinossauro cuja linhagem se encontra mais próxima à origem dos terópodes, apresenta algumas vértebras com espinhos neurais e centros sem pneumaticidade. Isso sugere que a pneumatização nos terópodes pode ter evoluído de forma diferente da observada nos saurópodes. Por outro lado, Rahonavis, um paraviano próximo aos raptores e das aves, apresenta uma pneumatização mais complexa, o que pode ter proporcionado vantagens em voos e escaladas.
Figura acima apresenta a tomografia computadorizada de vértebras cervicais de diversos terópodes. Note o aumento do volume e da complexidade da pneumaticidade desde os Ceratosauria até as Aves. Fonte: Aureliano et al. (2024).
Comparando com outros dinossauros terópodes representantes dos grandes grupos, é possível observar um aumento na pneumaticidade e na complexidade destas estruturas desde o Majungasaurus até as cegonhas de hoje em dia.
A compreensão da evolução do sistema de sacos aéreos nos terópodes tem implicações para a biologia evolutiva e para a paleontologia. Ela pode ajudar a elucidar como os dinossauros se adaptaram a diferentes ambientes e como essas adaptações influenciaram na sua sobrevivência. Além disso, futuros estudos sobre os primeiros terópodes vão poder fornecer mais informações sobre a evolução deste sistema.
Essa pesquisa foi financiada pela Fundação Norte-Rio-Grandense de Pesquisa e Cultura, e pela Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico.
Vídeo do nosso canal sobre esta pesquisa:
Bibliografia:
Aureliano, T., Almeida, W., Rasaona, M., & Ghilardi, A. M. 2024. The evolution of the air sac system in theropod dinosaurs: Evidence from the Upper Cretaceous of Madagascar. Journal of Anatomy. Link: https://doi.org/10.1111/joa.14113
Por Matheus P. dos Santos da Rocha & Cledston Matheus A. Macário
Quando falamos em Paleontologia, muitos a resumem como uma ciência meramente de descrição de aspectos morfológicos, como o simples trabalho de encontrar um osso, descreve-lo e, por sorte, dar nome a uma nova espécie. Porém, a Paleontologia vai muito além disso. Por meio dela, podemos especular sobre diversos aspectos da vida no passado. Até mesmo alguns cujas evidências, muitas vezes, são escassas no registro fossilífero. Um exemplo disso, seria encontrar uma resposta para a pergunta: como eram os olhos dos dinossauros não-avianos?
Como toda ciência, a paleontologia trabalha, inicialmente, com hipóteses, e essas, podem nos levar para linhas de raciocínio beeeem inusitadas, uma hora podemos estar debatendo sobre buracos negros e a extinção dos dinossauros e isso, mais à frente, pode terminar numa deliciosa (ou não) receita de macarrão com biscoito. A história de hoje começa com uma linhas de raciocínio inusitadas: ela parte de um grupo de insetos fósseis, os Kalligrammatidae…
O que são os Kalligrammatidae?
Chrysoperla carnea – Foto de Julia Stoess
Você já viu em algum jardim por aí pequenas bolinhas sustentadas por um fio bem fino, presas nas folhas das plantas? Se sim, com quase toda certeza você viu ovos de bicho-lixeiro. Pertencentes a uma ordem de insetos chamada Neuroptera, esses inofensivos (para os humanos) insetos são predadores vorazes de ovos de aranhas e outros invertebrados. Essa ordem inclui desde a formiga-leão, até coisas estranhas como os mantispídeos (que parecem uma mistura bizarra entre um marimbondo e um louva-a-deus).
Apesar de não serem um grupo muito comum nos dias de hoje, a representação fóssil deles é abundante. No Brasil, dados de um trabalho de revisão de 2018, dão conta que das 379 espécies de insetos descritos para a Formação Crato, da Bacia do Araripe, 76 são neurópteros, ou seja, 20% da diversidade de insetos da formação está em uma única ordem, que atualmente representa 0,6% das espécies de insetos viventes.
No meio de toda essa diversidade, os fósseis mais enigmáticos de Neuroptera são os da família Kalligrammatidae. O primeiro de Kalligrammatidae foi descrito por Johannes Walther, em 1904, com base num material quase completo, encontrado no calcário jurássico de Solnhofen (Alemanha) – aquele mesmo do Archaeopteryx. Desde então, diversas espécies de Kalligrammatidae foram encontradas em várias localidades, com destaque para os achados na China e nos âmbares birmaneses, ao norte de Mianmar.
Diversidade dos kalligrammatidae. a. & b. da Formação Crato (Brasil); c., d., e., f., g., h. & p. das Formações Jiulongshan ou Haifanggou (China); i., j., k., l., m. & n. da Formação Yixian (China); o. da Formação Karabastau (Cazaquistão).” – Imagem original por Julian Kiely (Editado).
Em 1997, o lendário paleontólogo Rafael G. Martins-Neto, descreveu, pela primeira vez, um Kalligrammatidae na Formação Crato, batizado de Makarkinia adamsi. De lá pra cá, outros trabalhos confirmaram a presença dessa família no Nordeste Brasileiro e, inclusive, descreveram novas espécies, sendo este, até hoje, o único lugar fora da Europa e Ásia a ter esses registros.
É uma sorte que esses animais ocorram em vários afloramentos do tipo lagerstätten (sítios com preservação excepcional) pelo mundo afora. A boa preservação dos fósseis permitiu notar rapidamente a semelhança dos kalligrammatídeos fósseis com as atuais borboletas e mariposas. Essa comparação não fica só por conta do formato, padrões de coloração e desenhos das asas, mas alguns espécimes bem preservados, principalmente em âmbar, mostram também a presença de uma “boca” modificada em um fino e comprido tubo chamado de probóscide, característica marcante das mariposas e borboletas (ambas pertencentes à ordem Lepidoptera). Mas isso aconteceu nos kalligrammatídeos num momento do tempo geológico em que as borboletas não existiam e as mariposas não eram tão abundantes e diversificadas como são hoje.
Fora do Brasil, alguns Kalligrammatidae chegam a ser apelidados de “giant lacewings” (crisopídeos gigantes) e isso chegou ao extremo em algumas espécies fósseis. Comparativamente, algumas espécies fósseis são enormes em relação aos seus irmãos ainda viventes. Estima-se que as espécies encontradas no Araripe, por exemplo, alcançavam entre 24 a 32 centímetros de envergadura!
A história dos “olhos” nas asas
Insetos grandes e chamativos podem virar comida facilmente, por isso, precisam ter alguma forma de se proteger da predação. Os kalligrammatídeos que viveram entre o Eojurássico ao Neocretáceo estavam dividindo espaço com lagartos, dinossauros avianos e não-avianos, pterossauros, entre outros predadores . Logo, teria que haver alguma forma deles não sucumbirem a seus colegas de habitat!
As mariposas e borboletas de hoje em dia têm algumas estratégias para evitar a predação. Desde projeções nas asas para desviar a atenção do predador, como as mariposas do gênero Actias, até mimetizar (imitar) folhas secas, tal qual Zaretis itys faz. Outra forma é ter “olhos”, ou melhor, ocelos em suas asas. Os ocelos são desenhos circulares que aparecem em diversos animais, especialmente nos lepidópteros. Esses círculos podem aparecer com 2 estratégias diferentes de uso:
Mycalesis patnia – Foto por L. Shyamal
A primeira é ter eles próximos às margens da asa, fazendo com que a atenção de um provável predador seja focada na ponta da asa e não no centro do corpo do organismo.
A outra é simplesmente aterrorizar! As mariposas da família Saturniidae e as borboletas-olho-de-coruja do gênero Caligo, por exemplo, fazem isso muito bem. Elas têm ocelos enormes no centro das asas, que imitam – algumas vezes de forma assustadora – os olhos de uma coruja, afastando assim qualquer predador que ouse atacá-las.
Caligo beltrao – Foto por Quartl
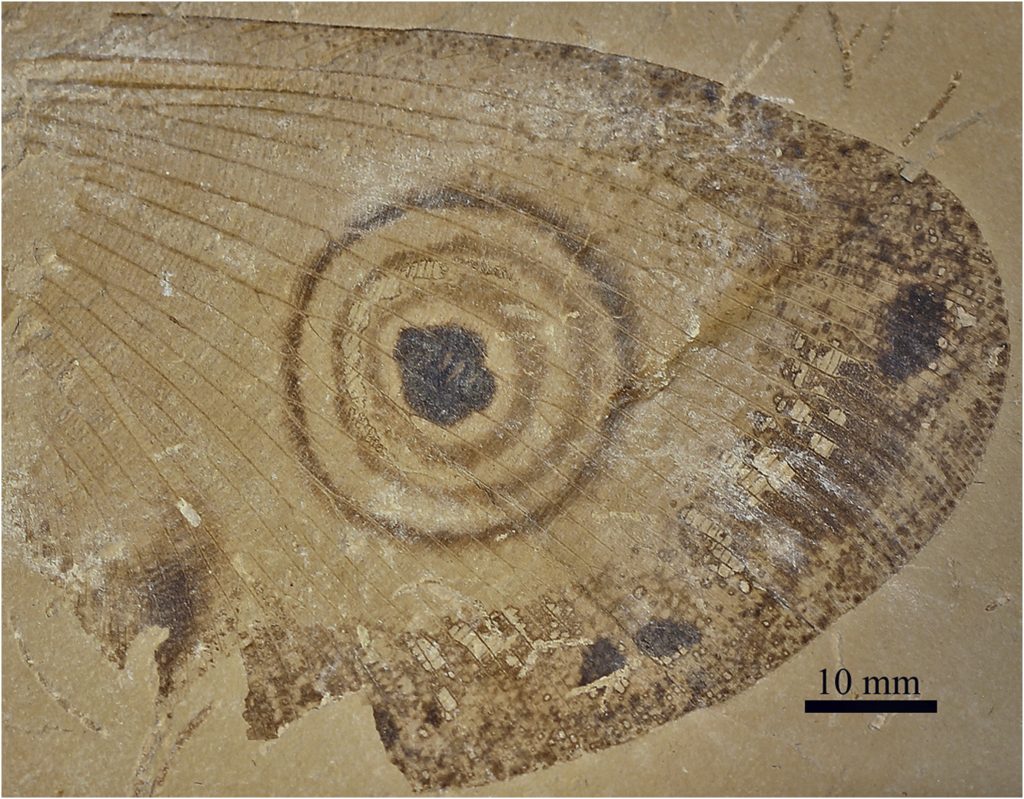
E é nesse ponto que queríamos chegar. Justamente essa segunda estratégia é atribuída a várias espécies fósseis de kalligramatídeos. Desde o primeiro espécime descrito, os ocelos gigantes estão presentes nas asas, e há trabalhos que descrevem e comparam os diversos formatos encontrados.
Makarkinia irmae – Imagem de Machado et al. (2021).
O que isso tem a ver com dinossauros?
Agora, chegou a hora que, ou vocês sairão desse blog nos chamando de loucos, ou terão o famoso “Mind Blow”. Vamos ao ponto principal: você já parou para pensar sobre o formato dos olhos dos dinossauros não-avianos? Essa é uma discussão complicada, pois o número de olhos de dinossauro preservados no registro fossilífero é: zero! Mas é uma curiosidade legítima querer saber essa informação, tanto que pode ser encontrado por aí, em fóruns pela internet, pessoas debatendo sobre essa questão.
Como esse tipo de material fóssil para dinossauros é inexistente, parte-se para a comparação com animais recentes, tanto seus parentes mais próximos ainda vivos, quanto possíveis análogos ecológicos. Mas existe ainda outra linha de raciocínio para se debater: não olhar para os dinossauros em si, mas para seus colegas de habitat e, no nosso caso especifico, os kalligramatídeos da Formação Crato.
A reação dos leitores daqui a alguns instantes, pelo menos, na expectativa dos autores…
Como já foi mencionado anteriormente, os “Giant Lacewings” poderiam ter se utilizado da segunda estratégia de uso dos ocelos: para assustar prováveis predadores, imitando os olhos de animais com os quais conviveram. Aí está o “pulo do gato”. Para um predador se assustar com os olhos desenhados nas asas das borboletas-olho-de-coruja é preciso que tenha um animal no mesmo habitat, que vá servir de gatilho (o “modelo” dos ocelos de Caligo, uma coruja, por exemplo: um predador assustador, que assuste o predador da Caligo). Mas há 120 milhões de anos não existiam corujas no Ceará, então…quem eram os modelos dos Kalligrammatidae do Crato?
Pantano do Crato – Arte de Olmagon.
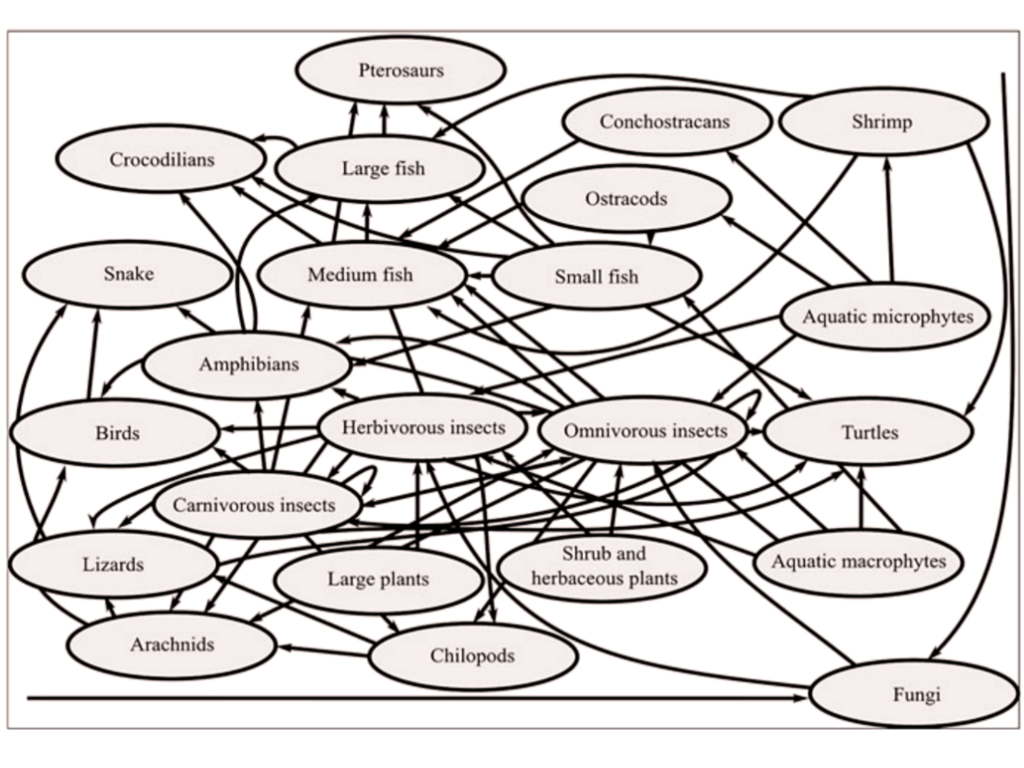
Existem dois principais suspeitos: pterossauros e dinossauros, mas vamos por partes. Pterossauros na Bacia do Araripe, segundo Mendes et al. (2020), eram majoritariamente piscívoros (comedores de peixes), com algumas exceções como Lacusovagus magnificens, que provavelmente vagava pelos pântanos da região para caçar anfíbios e outras pequenas presas. O trabalho de Mendes, inclusive, coloca os pterossauros como animais no topo da teia trófica da região na época.
“Teia trófica da fauna Cretácea do Araripe” – Mendel et al. (2020)
Mas se os pterossauros cearenses comiam peixes, majoritariamente, os possíveis predadores dos kalligramatídeos (outros insetos, anfíbios, pássaros, pequenos dinossauros, etc.) não estavam no cardápio deles, a priori. Por esse fator, seria compreensível a exclusão desses animais como possíveis modelos para os ocelos.
Escultura do Santanaraptor placidus do Museu Plácido Cidade Nuvens, de Santana do Cariri, CE.
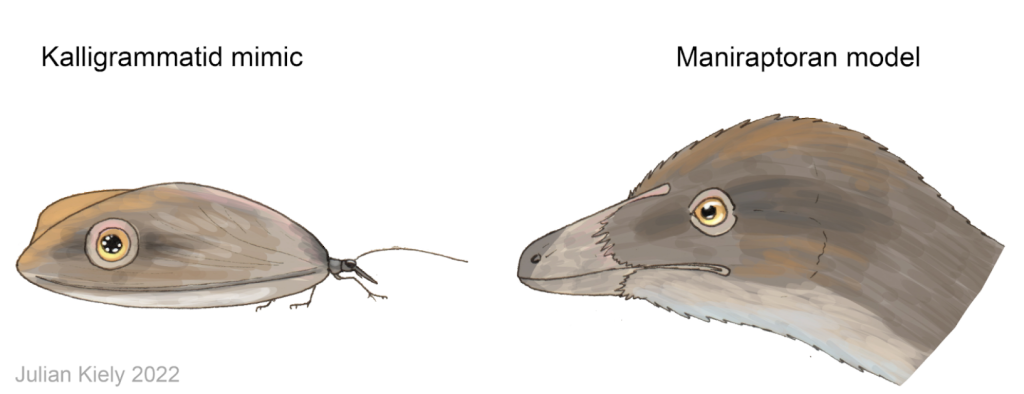
Já os dinossauros, por outro lado, são os candidatos perfeitos para esse quebra-cabeças ecológico. Animais como Aratasaurus museunacionali, Mirischia asymetrica (que, assim como “Ubirajara“, foi traficado para Alemanha #MirischiaBelongtoBR) e Santanaraptor placidus, ocupavam o nicho de predadores de médio a pequeno porte da região do Cariri. Como apontado por Julian Kiely, em seu artigo para o blog “Paleoflora”, a forma da asa dianteira na maioria das espécies de kalligramatídeos, e o grande tamanho dessas asas, correspondiam, aproximadamente, ao tamanho e a forma das cabeças de muitos pequenos dinossauros predadoresque conviviam com esses insetos (como as espécies mencionadas acima). Desta forma, poderíamos inferir que as pupilas dos dinossauros de médio a pequeno porte do Jurássico Superior e do Cretáceo Inferior, como os maniraptores (pelo menos), deveriam ser arredondadas, já que os ocelos de todos os kalligramatídeos conhecidos até então, possuem esse mesmo formato. O que se soma à evidência indireta parelela, que considera como base comparativa o formato da pupila dos dinossauros viventes, que são as aves.
Mimetismo de Kalligrammatidae a um Maniraptora – Imagem de Julian Kiely, 2022.
O poder da especulação
Alguns podem estar se perguntando: qual a importância de especular aspectos biológicos e evolutivos tão difíceis de se comprovar por meio do registro fossilífero? Muito da ciência começa com especulação. As descobertas científicas, em geral, nascem de hipótese de alguém. Um exemplo clássico foi a detecção das ondas gravitacionais em 2015, que haviam sido previstas por Albert Einstein em 1916.
Em homenagem à previsão de Meszaros, Tamisiocaris foi incluído em um novo clado denominado Cetiocaridae. Infelizmente, o nome deste clado não é mais considerado válido, de acordo com o Código Internacional de Nomenclatura Zoológica, por não existir nenhum gênero real chamado “Cetiocaris“, então foi formalmente substituído pelo nome Tamisiocarididae.
Reconstrução de Tamisiocaris – Arte de Rob Nicholls
Finalmentes
Com base em todos os argumentos supracitados (Alô, professores de redação!), podemos inferir que a hipótese levantada pode levar a especulações e trabalhos futuros que respondam às nossas dúvidas (isso claro, se esse post, neste humilde blog, chegar nas pessoas certas, e para isso seu compartilhamento é fundamental). Gostaríamos de agradecer a Julian Kiely do excelente blog “Paleoflora” pelo artigo que inspirou este, e que isso inspire a todos os nossos leitores a imaginar e especular dentro da ciência, lançar ideias, compartilhá-las, pois só assim a ciência cresce e prospera, com união e partilha.
Referências:
Martins-Neto, R. G. 1997. Neurópteros (Insecta, Planipennia) da Formação Santana (Cretáceo IInferior) Bacia do Araripe, Nordeste do Brasil. X – descrição de novos taxa (Chrysopidae, Babinskaiidae, Myrmeleontidae, Ascalaphidae e Psychopsidae). Revista Universidade Guarulhos , São Paulo, v. 2, n.4,. p. 68-83.
A espécie humana está na Terra há apenas 300 mil anos. Somos jovens nesse pequena planeta azul e dinâmico. Os dinossauros, por sua vez, estão por aqui há pelo menos 233 milhões de anos, desde o Período Triássico e, não custa lembrar, permanecem vivos até hoje na forma das aves. Esse grupo de animais tolerou e se adaptou a uma grande variedade de climas e mudanças dramáticas na configuração dos continentes ao longo do tempo. Por isso são um modelo excelente para estudarmos evolução biológica. Eles têm muito a nos ensinar sobre os segredos da sobrevivência.
Durante o auge do reinado dos dinossauros, na Era Mesozoica, o clima do nosso planeta era muito mais quente do que hoje. Uma das características que favoreceu este grupo de animais foi a evolução de sacos aéreos, um tipo de upgrade do sistema respiratório. Os sacos aéreos são estruturas conectadas aos pulmões, que se espalham por toda cavidade toráxica e abdominal desses animais, penetrando inclusive os ossos. Estão presentes nas aves atuais e não apenas tornam sua respiração mais eficiente, mas também ajudam a deixar os seus esqueletos mais leves, o que favorece, por exemplo, o voo. Apesar de muito característicos das aves, os sacos aéreos não são uma exclusividade dos delas. Eles também estavam presentes nos dinossauros não-avianos (todos os outros dinossauros, que não as aves) muito antes da evolução do voo.
Esquema mostrando os sacos aéreos em aves atuais. Fonte: https://pt.wikipedia.org/wiki/Sacos_a%C3%A9reos
Imagina-se que os sacos aéreos originalmente favoreceram os dinossauros por funcionarem como um sistema eficiente de captação de oxigênio e também por serem um sistema de refrigeração natural. Se você, hoje, fica ofegante fazendo exercícios no verão quente, saiba que os dinossauros eram (e são!) muito mais eficientes que você em captar oxigênio e se refrigerar. Não é à toa que eles saíram na frente na corrida evolutiva (enquanto nosso grupo, o dos mamíferos, ficou por quase 150 milhões de anos no banquinho de reservas evolutivo).
Já é bem sabido que dinossauros do Período Cretáceo, como o T. rex e alguns pescoçudos, como o Ibirania, tinham um extenso sistema de sacos aéreos pelo corpo. Inclusive, bem parecido com os das aves atuais. Só que a origem e evolução deste sistema tem sido um enigma por várias décadas. Será que os primeiros dinossauros, lá do período Triássico, já tinham sacos aéreos?
O que sabíamos era que a pneumaticidade do esqueleto relacionada a um sistema de sacos aéreos estava presente tanto em dinossauros derivados, ou seja, aqueles que viveram durante o Período Cretáceo, quanto em pterossauros, répteis voadores parentes próximos dos dinossauros. Ambos os grupos seguiram um caminho evolutivo independente a partir do Período Triássico. Uma explicação para a presença de sacos aéreos tanto em dinossauros quanto em pterossauros seria que a origem dessas estruturas se deu bem antes deles terem seguido seu caminho evolutivo independente, isto é, ainda em seus ancestrais.
Porém, a questão permaneceu em aberto. Faltavam estudos avaliando a presença dessas estruturas tanto em dinossauros mais antigos quanto em ancestrais dos pterossauros e dinossauros…
Para nossa sorte, o Brasil têm os fósseis dos mais antigos dinossauros e é aí que entra o estudo publicado agora em Dezembro de 2022 pelo nosso grupo de pesquisa, na revista Scientific Reports:
Para tentar solucionar este enigma, um grupo de pesquisadores brasileiros da Unicamp, UFRN, UFSCar e UFSM e um colaborador da Western University of Health Sciences, dos E.U.A., analisaram três fósseis de alguns dos mais antigos dinossauros do mundo, Buriolestes, Pampadromaeus e Gnathovorax, do Período Triássico do Rio Grande do Sul. Estes são alguns dos dinossauros mais antigos conhecidos até o momento, com 233 milhões de anos de idade!
Reconstrução do dinossauro herrerassaurídeo Gnathovorax. Arte por Márcio L. Castro.
Foi possível notar que os ossos da coluna vertebral (vértebras) desses animais apresentavam pequenos orifícios nas laterais. Sabemos que os sacos aéreos ingressam no esqueleto através de estruturas semelhantes a isso. Porém, os orifícios encontrados eram muito pequenos, o que talvez indicasse uma outra função.
Realizamos, então, tomografias de alta resolução (micro-tomografias) para investigar a estrutura interna dos fósseis. A análise revelou uma arquitetura bastante densa nas vértebras desses animais, bem diferente do que conhecemos em esqueletos permeados por sacos aéreos de dinossauros que viveram no Cretáceo ou mesmo as Aves. Porém, Buriolestes e Pampadromaeus mostraram uma vascularidade mais complexa no interior das vértebras, do que Gnathovorax. Uma vascularidade mais desenvolvida pode ter servido de alicerce para o surgimento das estruturas pneumáticas conhecidas como câmaras e camelas, típicas da invasão das vértebras por sacos aéreos.
Reconstrução do dinossauro Pampadromaeus. Arte por Márcio L. Castro.
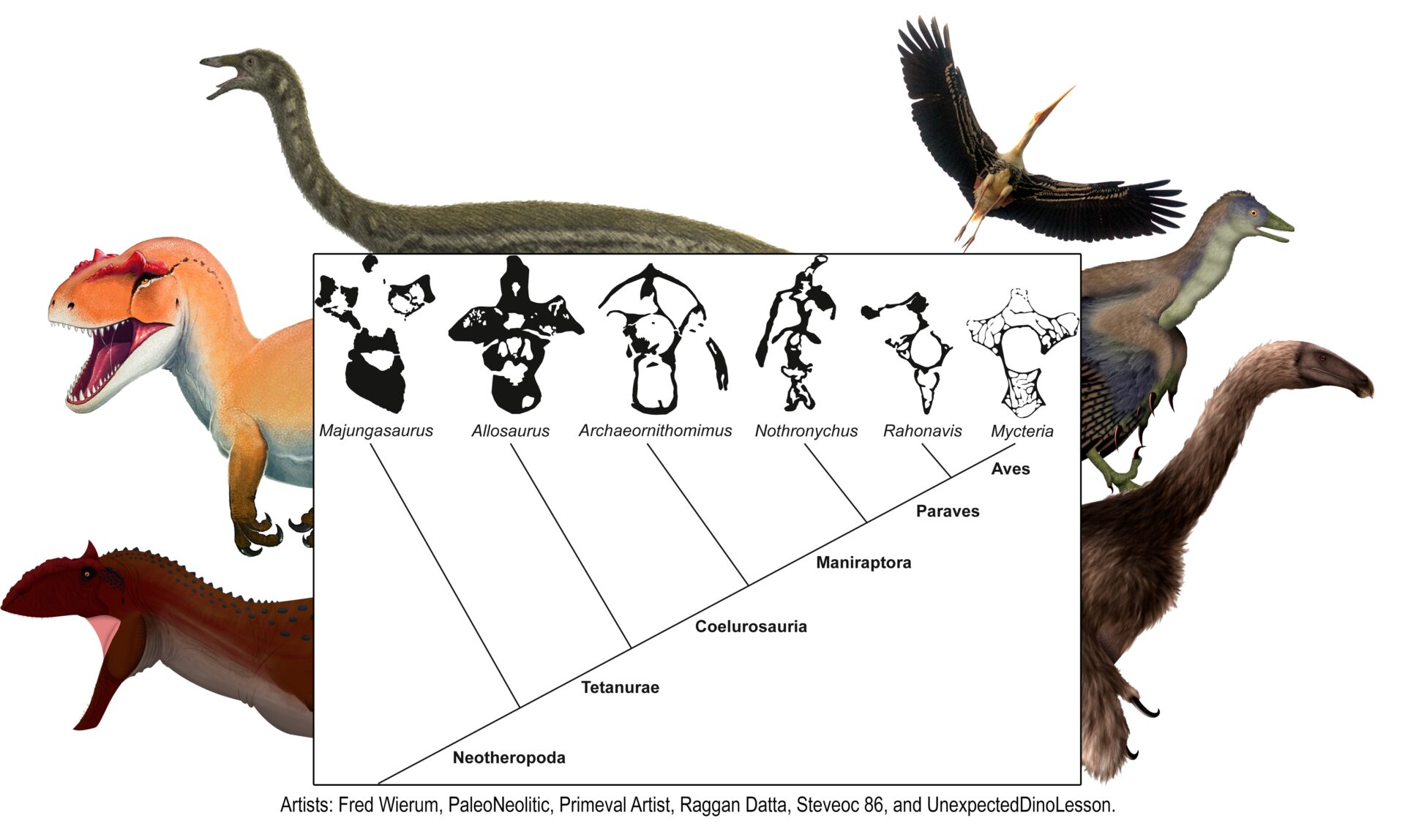
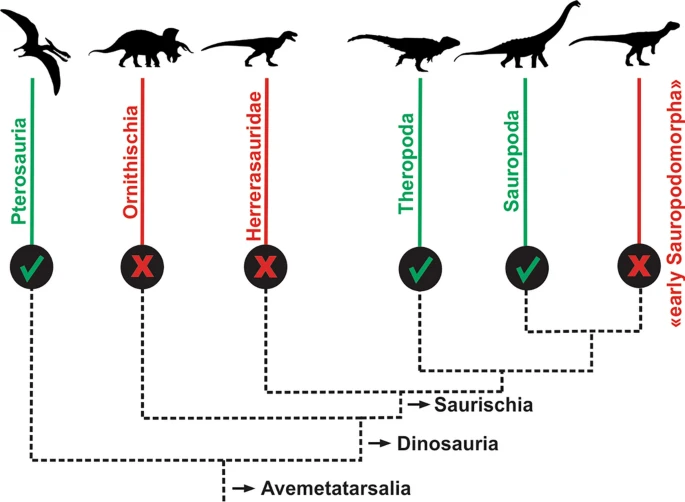
A ausência de pneumaticidade no esqueleto pós-craniano desses dinossauros mais antigos contradiz a hipótese de que os sacos aéreos invasivos presentes em dinossauros e pterossauros são homólogos, ou seja, de que teriam surgido no ancestral comum desses animais. Isso indica que a pneumaticidade óssea associada à sacos aéreos evoluiu pelo menos três vezes independentemente em Avemetatarsalia, grupo que inclui dinossauros, pterossauros e seus parentes. Ou seja, evoluiu de forma independente em pterossauros, dinossauros terópodes (grupo dos dinossauros carnívoros) e sauropodomorfos (grupo dos dinossauros pescoçudos).
Uma árvore simplificada dos dinossauros e seus parentes mostrando a evolução independente dos sacos aéreos em pterossauros, dinossauros terópodes e sauropodomorfos.
Essa descoberta muda a forma como compreendíamos os dinossauros e seus parentes. Passo a passo estamos entendendo melhor a sua evolução e o segredo do seu sucesso. É possível que algum fator ambiental tenha sido o gatilho para a evolução desse sistema sacos aéreos em diferentes grupos de avemetatarsalianos, mas isso são cenas para os próximos capítulos!
Gostaríamos de agradecer as agências de fomento que tornaram possível esta pesquisa: o CNPq, a FAPESP e a FAPERGS.
Acesse o artigo completo: Aureliano et al. 2022. The absence of an invasive air sac system in the earliest dinosaurs suggests multiple origins of vertebral pneumaticity. Scientific Reports. https://www.nature.com/articles/s41598-022-25067-8
Os maiores animais a caminharem em terra firme foram os dinossauros saurópodes, apelidados de pescoçudos. Algumas espécies de pescoçudos, como o Argentinosaurus ou o Patagotitan, encontrados na Argentina, podiam ultrapassar 30 metros de comprimento. Verdadeiros colossos capazes de fazer a terra tremer! Mas nem todos os saurópodes eram assim… Existiram centenas de espécies desses dinossauros em quase todos os continentes e, apesar da maioria ser conhecida pelo seu grande tamanho, algumas formas adotaram uma tendência contrária. Existem alguns casos de pescoçudos anões, formas com a altura de um cavalo ou de um camelo, como Magyarosaurus ou Europasaurus, encontrados em ambientes de ilhas antigas. Via de regra, essas formas anãs são encontradas em ambientes de ilhas, pois devido a restrição de área e recursos, a miniaturização do corpo pode ser uma vantagem. Porém, para nossa surpresa, fósseis de uma nova espécie de dinossauro pescoçudo anão foram encontradas aqui no interior do Brasil, em um lugar que esteve bem longe do mar durante toda a Era dos Dinossauros. Essa espécie de dinossauro foi descoberta na cidade de Ibirá, no interior de São Paulo, e se tornou uma das menores espécies de dinossauros pescoçudos conhecidas do mundo!
Reconstituição da nova espécie de dinossauro anão de Ibirá. Arte por Matheus Gadelha.
Por mais de 15 anos o Prof. Marcelo Fernandes (UFSCar) e seu grupo de pesquisa, eu inclusa, têm coletado fósseis no Noroeste Paulista, em uma localidade onde são encontrados abundantes fósseis de dinossauros. As rochas e fósseis dessa localidade datam do Período Cretáceo e têm aproximadamente 80 milhões de anos. Dentre os fósseis recuperados estão restos de dinossauros carnívoros, crocodilos, tartarugas e vários outros animais da “Era dos Dinossauros”. Muitos restos de dinossauros herbívoros foram encontrados na localidade, mas até o momento nenhuma espécie de pescoçudo havia sido nomeada para a região.
Eu procurando por fósseis no sítio onde foram encontrados fósseis do pequeno pescoçudo em Ibirá, SP. Foto por Tito Aureliano.
Fui eu quem trabalhou pela primeira vez, durante a minha graduação, com os fósseis do pequeno dinossauro pescoçudo de Ibirá. Àquela época, o dinossauro não ganhou nome, mas foi reconhecido como diferente das outras espécies descritas para o Brasil até então. Muito tempo se passou, mais fósseis desse pequeno dinossauro foram encontrados e, finalmente, alguns anos atrás, a missão de liderar a descrição da espécie desse misterioso dinossauro nanico foi dada ao paleontólogo Bruno Navarro, atualmente estudante de doutorado no Museu de Zoologia da USP, e especialista em dinossauros saurópodes. Bruno, Marcelo e eu contamos com a ajuda de uma excelente equipe de colaboradores nesse processo e, no último dia 15 de setembro, apresentamos formalmente essa nova espécie de dinossauro ao mundo científico.
O colega Bruno Navarro em Ibirá, SP, procurando por fósseis. Foto do arquivo pessoal de Bruno.
Comparando os fósseis do pequeno dinossauro de Ibirá com materiais de outros animais do mesmo grupo encontrados no Brasil e no mundo, foi possível concluir que ele pertencia à família dos saltassauros, um grupo de titanossauros que inclui algumas espécies de já de tamanho bastante reduzido. Além disso, o pequeno dinossauro de Ibirá apresentava várias características únicas, não compartilhadas com seus parentes mais próximos, logo, uma nova espécie poderia ser batizada. O nome escolhido foi Ibirania parva.Ibirania é a junção das palavras Ibirá – cidade onde a espécie foi encontrada – e ania que em grego significa “caminhante, peregrino”. Já parva é o latim para ‘pequeno’. Como a palavra Ibirá vem do Tupi para “Árvore” – é possível traduzir o nome desse dinossauro como “o pequeno peregrino das árvores”.
Reconstituição artística de Ibirania parva por Hugo Cafasso.
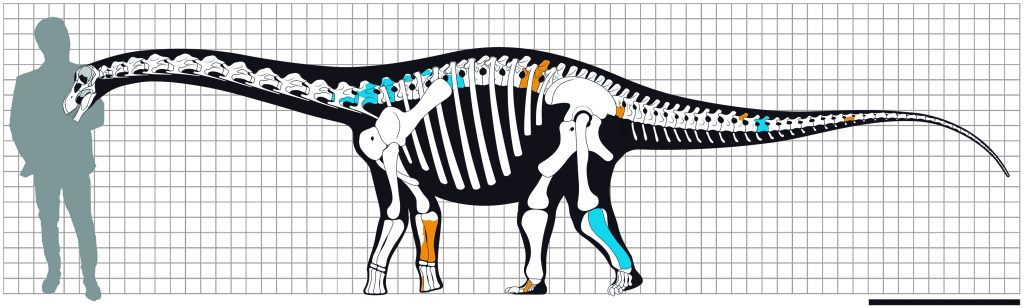
Desde o princípio era possível notar que os fósseis desse pescoçudo de Ibirá eram muito pequenos quando comparado a outros titanossauros, mas ao estimar o tamanho aproximado de um dos espécimes analisados, nos surpreendemos. Ele teria entre 5 e 6 metros de comprimento e seria da altura de uma vaca, o que o colocaria entre as menores espécies de saurópodes já descritas do mundo! Para checar se o tamanho reduzido seria porque o espécime era apenas um jovem quando morreu, resolvemos analisar o tecido ósseo fossilizado do dinossauro ao microscópio. Essas amostras foram analisadas pelo paleontólogo Tito Aureliano, atualmente estudante de doutorado da Unicamp. A partir da análise do tecido ósseo foi possível concluir que Ibirania realmente era uma espécie de titanossauro anão, já que os fósseis pertenciam a um animal adulto no momento de sua morte, ou seja, ele não cresceria mais ao longo de sua vida.
Tamanho estimado de Ibirania parva comparado a um humano de 1,80m. Em destaque as partes descobertas do esqueleto.
Vértebra dorsal de Ibirania parva. Imagem de Navarro et al. (2022). Escala = 10cm.
No interior de São Paulo, durante o final do Período Cretáceo, há 80 milhões de anos, caminharam muitos dinossauros pescoçudos de grande tamanho, e até gigantes, como o Austroposeidon. Mas havia algo de especial na região de Ibirá, que favoreceu a existência de pescoçudos nanicos. Diferente de outros anões que viviam em ilhas tropicais onde hoje é a Europa, como Magyarosaurus ou Europasaurus, Ibirania vivia no interior do Brasil, em um ambiente semi-árido com períodos chuvosos intercalados por secas intensas. Foi esse ambiente hostil, com recursos limitados periodicamente, que selecionou esses pequenos dinossaurinhos herbívoros, que ao invés de migrar, provavelmente permaneciam residentes na região.
Ibirania é a primeira espécie comprovadamente anã das Américas e viveu em um contexto muito diferente dos outros dinossauros pescoçudos anões já encontrados. Ela acrescenta novas informações sobre a evolução dos titanossauros e também sobre a ocorrência de nanismo em dinossauros saurópodes. Ibirania recebeu o apelido carinhoso de “Bilbo”, em referência ao hobbit de “O Senhor dos Anéis”, por ser um nanico entre gigantes. Se você quiser saber todas as descobertas que este ‘dinossauro-Hobbit’ já forneceu, assista à playlist: https://www.youtube.com/watch?v=_kH96sPGjfg&list=PLHPifkNwYyYYNFP-wvUXNti7NGkfNQ8hz.
O estudo foi publicado na revista Ameghiniana e pode ser acessado AQUI.
Assista também ao vídeo de divulgação:
Referência:
A. Navarro, B., M. Ghilardi, A. ., Aureliano, T., Díez Díaz, V., N. Bandeira, K. L., S. Cattaruzzi, A. G., V. Iori, F., M. Martine, A., B. Carvalho, A., Anelli, L. E., A. Fernandes, M., & Zaher, H. (2022). A NEW NANOID TITANOSAUR (DINOSAURIA: SAUROPODA) FROM THE UPPER CRETACEOUS OF BRAZIL. Ameghiniana, 59(5), 317-354. https://doi.org/10.5710/AMGH.25.08.2022.3477
Nas últimas postagens você teve a oportunidade de conhecer alguns grandes cetáceos fósseis predadores, como Basilosaurus isise Ankylorhiza tiedemani. Mas não só de caçadas sanguinolentas vivem esses animais. Bom, isso se você não for um cardume de lulas, peixes ou crustáceos...
Na postagem de hoje, vamos contar sobre como as baleias foram por um caminho um pouco diferente e se tornaram alguns dos maiores seres viventes do planeta, alimentando-se por filtração. Caso você seja parte do cardápio, é bom por sebo nas canelas, ou melhor, nas nadadeiras, porque a fome aqui é gigantesca.
Um balaio de baleia
Antes de qualquer coisa, é importante entender quem são as baleias filtradoras:
Em inglês, o termo whale(baleia)pode ser usado popularmente para se referir a qualquer espécie de cetáceo, seja este pequeno ou grande, com dentes ou não. Porém, também existe uma palavra para se referir especificamente às baleias filtradoras : baleen (algo como “barbas” ou “barbatanas”, em português).
“Baleen” refere-se particularmente aos Mysticeti ou “misticetos“, no bom português. Misticetos são os cetáceos dos grupos Balaenopteridae, Balaenidae, Eschrichtiidae, Neobalaeninae e alguns outros grupos fósseis. Estes grupos possuem barbatanas (ou barbas) no lugar dos dentes, que servem para filtração, e é por isso, que esses cetáceos são chamados de “baleias filtradoras” ou “baleias de barbatana” aqui no Brasil.
Também há nomes populares específicos para diferenciar certos grupos. Rorqual, por exemplo, refere-se somente às baleias da família Balaenopteridae, que incluem baleias-azuis, baleias-fin, baleias-sei, baleias-de-bryde, baleias-de-rice, baleias-minke e baleias-jubarte. Baleias verdadeiras, por sua vez, é o nome usado para denominar as Balaenidae, representas pelas baleias-francas-austrais, baleias-francas-do-atlântico-norte, baleias-francas-do-pacífico, e as baleias-da-groenlândia.
As outras duas famílias, Eschrichtiidae e Neobalaeninae, não ocorrem no Brasil, e normalmente são chamadas pelos nomes das espécies que as representam: baleia-cinzenta (Eschrichtiidae) e a rara baleia-franca-pigméia (Neobalaeninae).
Algumas espécies de baleias filtradoras (CASTRO & HUBER, 2012).
Modos à mesa
Cada grupo de baleia filtradora possui características anatômicas diferentes que refletem modos únicos de filtrar a água para se alimentar. Porém, pelo menos uma coisa é comum entre elas: as barbatanas, estruturas compridas e enfileiradas, feitas de queratina (mesmo material que forma nossos cabelos e unhas), presentes na boca desses animais. Tais barbatanas acabaram por substituir os dentes dos misticetos ao longo de sua evolução.
As barbatanas funcionam como uma escumadeira ou peneira, que permite que a água abocanhada durante a alimentação seja expulsa da boca, com a ajuda da língua, e que o alimento fique preso enquanto a água sai.
As rorquais possuem pregas na região ventral, que se expandem como o papo de um pelicano, permitindo com que elas possam abocanhar (engolfar) grandes quantidades de água com cardumes inteiros de pequenos peixes ou krill (um tipo de camarãozinho). Já as baleias-verdadeiras, são mais corpulentas, não possuem pregas ventrais e a sua boca é em forma de arco. Elas basicamente se alimentam filtrando a água enquanto nadam com a boca aberta.
As barbatanas das baleias-verdadeiras podem ter entre 2 metros de altura, até 5,2 metros de altura!
As baleias-cinzentas, por sua vez, abocanham a areia do fundo marinho filtrando-a em busca de crustáceos enterrados. Elas possuem sulcos na garganta em vez de pregas ventrais. As baleias-franca-pigméia, por fim, possuem uma mistura de características: boca em forma de arco como a das baleias verdadeiras, sulcos ventrais na garganta como as baleias-cinzentas e estilo de alimentação como as rorquais.
Diferenças entre as formas de filtração das rorquais (baleia-azul) e das baleias verdadeiras (baleia-franca). A água entra na boca e depois é expulsa com a ajuda da língua ao pressionar o céu da boca, fazendo a água passar pelas barbatanas (CASTRO & HUBER, 2012).
Mas sempre foi assim? Como será que as barbatanas surgiram? As formas de alimentação sempre foram as mesmas desde o começo da evolução dos misticetos? Como as espécies transicionais se alimentavam? Essas são perguntas que o registro fossilífero pode ajudar a responder…
Comendo com hashi, os “palitinhos japoneses“
Os basilossaurídeos foram os primeiros cetáceos totalmente aquáticos. Eles eram grandes caçadores, inclusive de outros cetáceos, e tinham a boca cheia de dentes diferentões, como já citamos AQUI. A pergunta inevitável é: como essas “baleias primitivas” foram de caçadoras dentadas para as banguelas filtradoras que temos na atualidade? Bom, alguns achados fósseis das últimas décadas possibilitaram entender parte desse processo:
Em 2016, Felix G. Marx junto com outros colegas, analisaram estranhas ranhuras encontradas em dentes fósseis de um grupo de baleias “primitivas” chamadas de Aetiocetidae, comuns no Oligoceno.
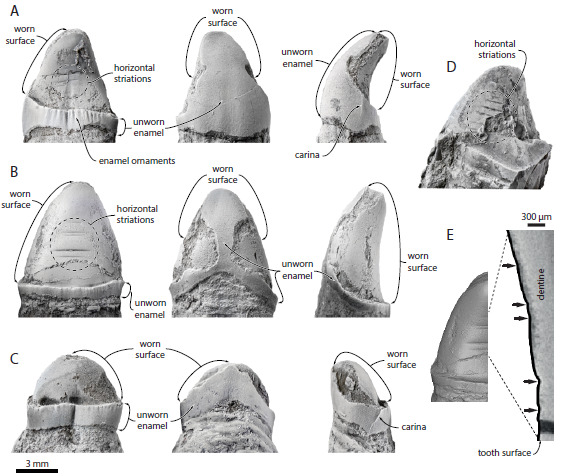
Crânio de aetiocetídeo (MARX et al., 2016).
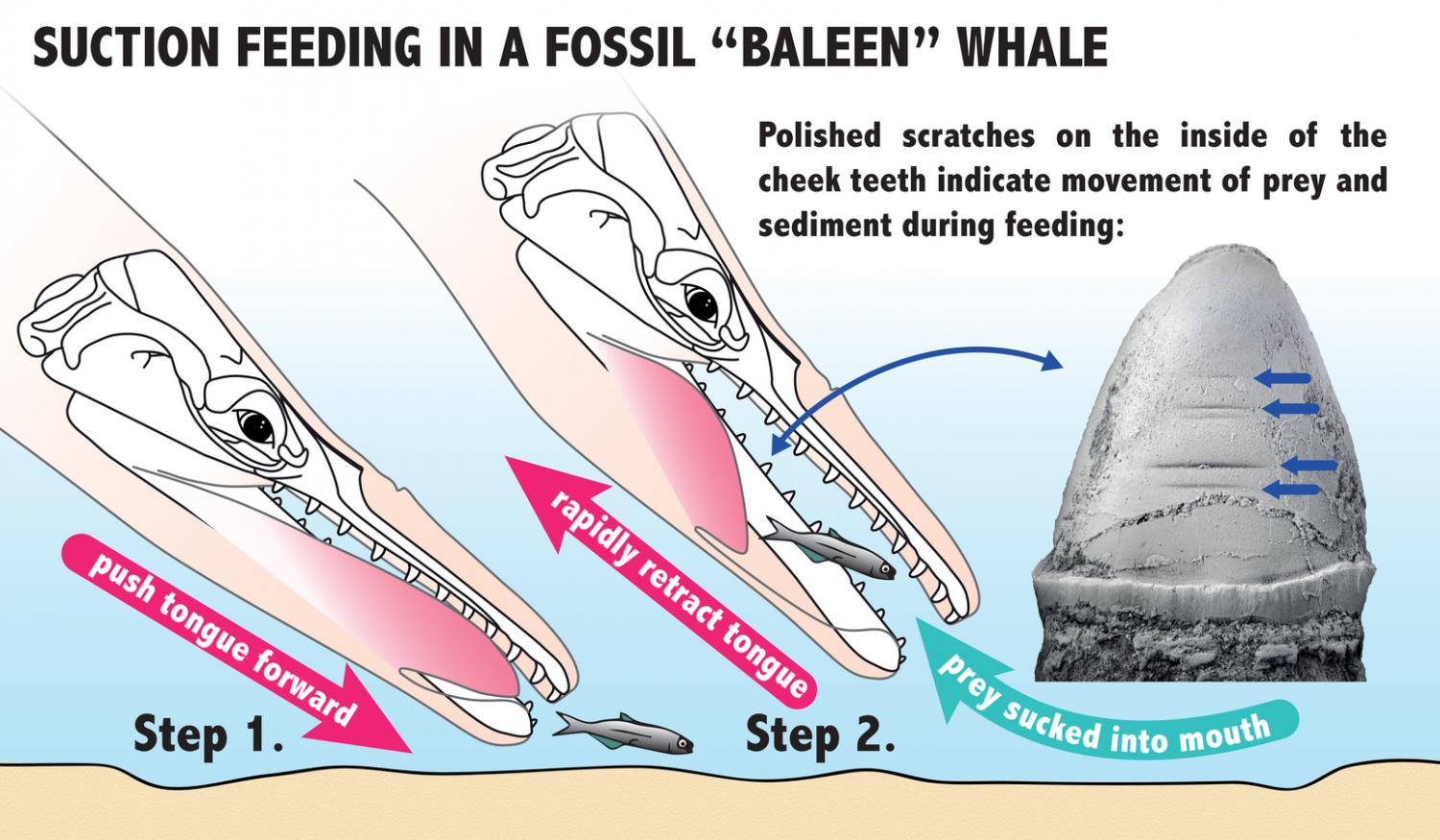
Eles analisaram um fóssil específico, encontrado em rochas da região de Washington, EUA. O espécime, datado do Oligoceno Superior demonstrou afinidade aos misticetos, mas possuía um padrão de desgaste nos dentes muito diferente, não compatível com a presença de barbatanas. Felix e colegas, depois de muitas análises, associaram os desgastes horizontais nos dentes a uma alimentação por sucção, sugerindo que as barbatanas dos misticetos teriam surgido mais tarde, próximo à origem das baleias filtradoras modernas. O que isso significa? Que antes de filtrar, os misticetos provavelmente teriam se alimentado sugando o alimento, como se faz quando comemos macarrão com hashi!
Figura 5. Padrões de desgaste em dentes, sugerindo alimentação por sucção em um aetiocetídeo (MARX et al., 2016).
As ranhuras horizontais observadas nos dentes foram comparadas com outros animais que também se alimentam por sucção na atualidade, como belugas, algumas baleias-bicudas, morsas e certas espécies de focas. Pelo padrão de desgaste, a língua deveria funcionar como um pistão, fazendo pressão para que a água e a(s) presa(s) fossem sugados rapidamente para dentro da boca, o que deveria causar estes desgastes.
Etapas de sucção mostrando a língua de um Aetiocetidae funcionando como um pistão. Ilustração por David Hocking.
Também foram observadas marcas em alguns dentes, relacionadas com a expulsão da água e sedimentos que eventualmente eram sugados junto com o alimento. Os Aetiocetidae deveriam ficar com a boca meio aberta enquanto expulsavam tudo aquilo que não era alimento. Essas marcas são semelhantes às encontradas em certas espécies de mamíferos marinhos citados anteriormente, que também se alimentam por sucção.
Mas e agora? Onde as barbatanas aparecem nessa história, se os primeiros misticetos ainda tinham dentes e sugavam a comida, ao invés de morder tudo que passava pela frente? Mais uma vez, o registro fossilífero pode nos ajudar a entender essa história, mas agora vamos contar com a ajuda de análises em animais atuais para entender outra parte dessa saga evolutiva única.
Comendo com garfo e colher
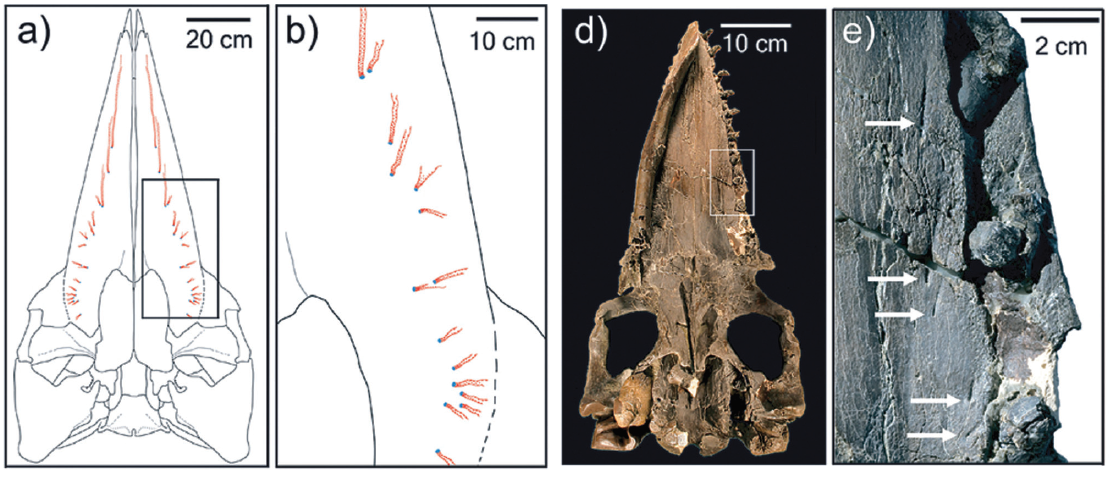
Alguns autores sustentam que as barbatanas teriam aparecido em alguns misticetos extintos que ainda possuíam dentes. Eles descrevem que essa teria sido uma “fase de transição” evolutiva, em que esses cetáceos teriam exibido uma alimentação mista: de predadores ativo por sucção, que também seriam capazes de realizar filtragem. Achados fósseis do Oligoceno Superior, publicados em 2008, por Thomas A. Deméré e colegas, permitiram o reconhecimento de marcas no céu da boca de espécies fósseis de Aetiocetidae, que indicariam uma alimentação por filtração.
Aetiocetus weltoni, um Aetiocetidae mostrando a hipótese de ocorrência simultânea de dentes e barbatanas (Ilustração de Carl Buell) (DEMÉRÉ et al., 2008).
O material descrito por Thomas pertence a uma espécie denominada Aetiocetus weltoni. Nos fósseis foram encontradas ranhuras de inervação no palato (céu da boca) semelhantes àquelas observadas nas baleias filtradoras atuais. As inervações saem da região dos alvéolos dentários (local onde se encaixam os dentes) exatamente como nos misticetos atuais. Segundo os autores, isso seria uma evidência de que a espécie fóssil teria tido barbatanas.
Palato de baleia atual sem dentes, mostrando os sulcos de inervações (a-b) e palato de Aetiocetus weltoni mostrando os sulcos de inervações junto com a dentição (MARX et al., 2016).
Inicialmente achava-se que as barbatanas poderiam ter surgido a partir de estruturas rígidas no céu da boca dos misticetos basais, chamadas de “cristas córneas palatinas”, também presentes nos artiodáctilos (cabras, bois, camelos, hipopótamos, etc.), grupo no qual os cetáceos são aparentados. Porém, as inervações encontradas nos fósseis observados por Thomas e colegas reforçaria outra hipótese.
Thomas e colegas observaram fetos de baleias filtradoras atuais e notaram que, apesar de “banguelas” quando adultas, a sua sequência de dentes se desenvolve em sua fase fetal. O seu crescimento é que é interrompido pela ação de alguns genes. Quando tais genes se ativam, os dentes são reabsorvidos pelo organismo, mantendo somente as inervações no palato geradas durante o seu crescimento inicial.
Vista lateral de um feto baleia-fin (Balaenoptera physalus) com corte mostrando botões de dente na mandíbula superior (MARX et al., 2016).
Durante a reabsorção dentária, queratina é secretada e é isso que forma as placas de barbatana no lugar dos dentes reabsorvidos. Essas placas crescem constantemente enquanto o filhote se desenvolve e, ao longo da vida, são desgastadas e desfiadas, ficando com a aparência que conhecemos.
A ordem dos talheres
Alguns autores, todavia, alertam: assim como a origem das penas em dinossauros não-avianos não marca a origem do vôo, o aparecimento dos canais de inervação no palaro dos misticetos não necessariamente indicaria a presença de barbatanas. Em vez disso, esses sulcos inervados em alguns aetiocetídeos fóssei poderiam, por exemplo, ter fornecido condições anatômicas e fisiológicas iniciais para o aparecimento posterior das barbatanas.
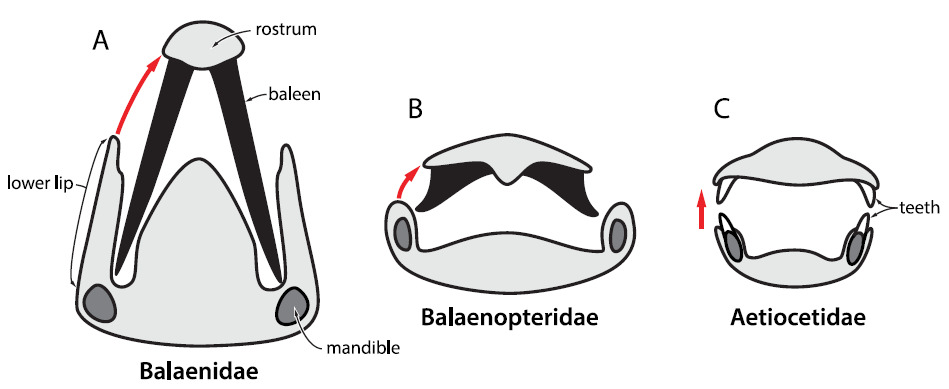
Felix G. Marx e colegas, em seu trabalho de 2016, apontam que a presença de dentes alternados, como os observados no fóssil descrito por Thomas e colaboradores em 2008, acabariam danificando as barbatanas, se essas estivessem presentes. Os mesmos autores também observam que a mandíbula das baleias filtradoras atuais é mais larga, o que permite com que as barbatanas não sejam danificadas durante o fechamento da boca do animal. Além disso, misticetos atuais também possuem uma adaptação muscular especial que permite com que a mandíbula rotacione levemente durante a oclusão, o que também dificulta o dano às barbatanas. Em aetiocetídeos, por sua vez, o tamanho da mandíbula e sua forma de abertura indicam que, qualquer barbatana, se presente, seria danificada durante o fechamento da boca.
Esquema de fechamento da boca e a acomodação das barbatanas nas baleias verdadeiras (Balaenidae) e nas rorquais (Balaenopteridae) em comparação com o fechamento da boca dos Aetiocetidae possivelmente sem barbatanas (MARX et al., 2016).
Felix e demais colegas até tentam dar uma chance ao modelo de barbatanas+dentes. Eles sugerem um cenário em que as barbatanas poderiam estar entre os dentes do animal e que, com o fechamento da boca, elas se dobrariam para dentro, similar ao que ocorre com as baleias-da-groenlândia. Todavia, ainda sim, a presença dos dentes alternados atrapalharia o dobramento e danificaria significativamente as estruturas. Os autores concluem, que as barbatanas não teriam surgido dessa forma. Talvez elas tenham surgido em outros grupos que tivessem uma mandíbula mais larga e/ou uma dentição reduzida ou inexistente.
Um jeito alternativo de comer
A capacidade de gerar sucção é fundamental para a maioria dos vertebrados aquáticos e é amplamente observada entre os mamíferos marinhos atuais. No entanto, até o trabalho de Felix e colegas, ela raramente havia sido associada à evolução dos misticetos. A sucção é muito útil na alimentação subaquática, pois facilita o transporte do alimento até o fundo da boca, onde ele será deglutido. Essa forma de adquirir alimentos provavelmente já estava presente em cetáceos basais, muito antes dos misticetos, mas esse é um comportamento relativamente difícil de se interpretar por meio de fósseis.
O uso de sucção na alimentação e provável ausência de barbatanas nos misticetos basais sugeriria um modelo diferente de evolução da alimentação por filtragem em baleias:
(1) Misticetos basais, incluindo aetiocetídeos, tinham tanto a dentição funcional, quanto a habilidade de usar sucção, herdada de cetáceos anteriores;
(2) A água ingerida por eles, como resultado da sucção, era expelida fundamentalmente com auxílio dos dentes;
(3) algum grupo de misticeto ancestral aprimorou suas capacidades de sucção ao longo do tempo, com o desenvolvimento de mandíbulas mais largas e tecidos moles associados (“calos” ou dobras na gengiva, lábios expandidos, etc.). O aperfeiçoamento desta capacidade teria favorecido a perda da dentição com o tempo;
(4) Com a perda da dentição, as barbatanas teriam evoluído e, gradualmente, sido selecionadas.
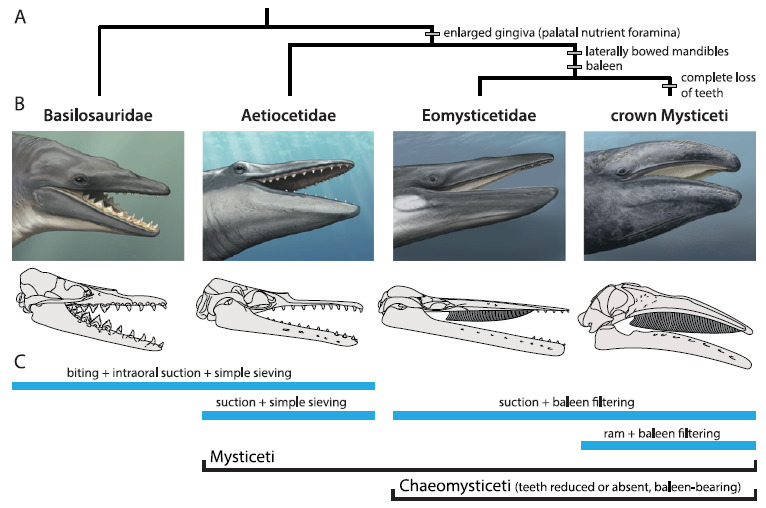
Esquema evolutivo das baleias filtradoras. (MARX et al., 2016).
Este cenário seria mais plausível, pois exclui problemas potenciais de interferência entre uma dentição ativa e barbatanas. Ele explica também como os dentes poderiam ter sido perdidos, sem afetar o sucesso da alimentação. Além disso, ele está de acordo com a evidência observada no desenvolvimento de fetos de misticetosatuais, que mostra que o crescimento das barbatanas só se dá quando os dentes são reabsorvidos.
A sucção seguida de filtragem é uma forma de alimentação que pode limitar bastante o tamanho máximo da presa a ser capturada. O aperfeiçoamento dos tecidos moles associados à dentição, em especial a gengiva, é uma forma de lidar com este problema. Primeiro, um mecanismo para prensar a presa deve ter surgido nos misticetos basais e, depois disso, um aparato apropriado de filtragem – isto é, as barbatanas.
Uma adaptação observada nos botos-de-dall (Phocoenoides dalli), espécie vivente de cetáceo, pode ajudar a imaginar como o processo teria acontecido. Esses cetáceos odontocetos (não diretamente aparentados às baleias filtradoras) têm dentes rudimentares e possuem em sua boca, também, pequenos “dentes na gengiva”, semelhantes aos brotos iniciais das barbatanas dos misticetos. Essas estruturas ajudam os botos-de-dall a capturarem suas presas. Pode ser que essa adaptação seja uma pista de como se deu a perda gradual da dentição em misticetos e posterior surgimento das barbatanas.
A origem das barbatanas, adaptação chave das baleias filtradoras modernas (Mysticeti), marca uma transição profunda e única na história evolutiva dos vertebrados. Hoje, ela é um pouco melhor compreendida, porém, como vocês viram, ainda restam detalhes a serem desvendados sobre como esta belíssima crônica evolutiva se desenrolou. A compreensão da evolução dos “modos à mesa” das baleias pode estar, não só nos fósseis, mas em uma combinação entre estudos paleontológicos, genéticos e do desenvolvimento.
Referências:
CASTRO, Peter; HUBER, Michael E. Biologia marinha. AMGH Editora, 2012.
DEMÉRÉ, Thomas A.; MCGOWEN, Michael R.; BERTA, Annalisa; GATESY, John. Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Systematic biology, v. 57, n. 1, p. 15-37, 2008.
MARX, Felix G.; HOCKING, David P.; PARK, Travis; ZIEGLER, Tim; EVANS, Alistair R.; FITZGERALD, Erich M. G. Suction feeding preceded filtering in baleen whale evolution. Memoirs of Museum Victoria, v. 75, 2016.