>
Quem já não assistiu o clássico “Querida, encolhi as crianças”? Para àqueles que ainda não viram, a aventura trata de um cientista que inventa uma máquina de “miniaturizar” objetos e acaba encolhendo seus filhos. Pequeninos e perdidos no jardim de casa, eles se deparam com abelhas, borboletas e formigas gigantes, que, dada a situação, parecem bichos aterrorizantes. Pois bem, se pudéssemos voltar no tempo alguns milhares ou milhões de anos, poderíamos vivenciar de fato esse cenário fictício, no entanto, sem precisar da famosa máquina encolhedora do cientista maluco…
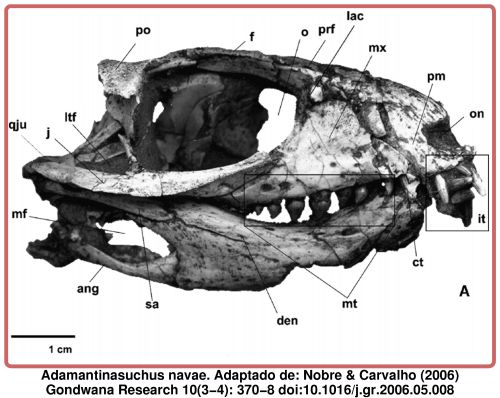
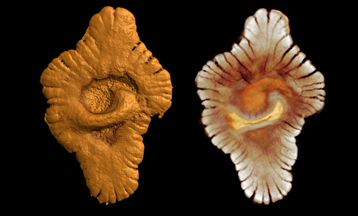
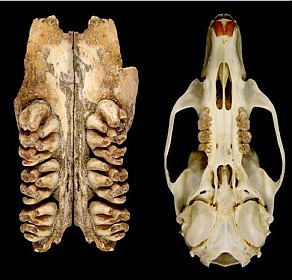
Pesquisadores encontraram recentemente uma ossada do que teria sido o maior rato já encontrado até hoje. Sim, um rato enorme. O animal, que se encaixa no gênero Coryphomys, e tem aproximadamente 1,5 mil anos, habitava as regiões do Timor Leste, na Ásia. O mais interessante – não se assustem – é que o tal bicho podia chegar a 6 kg, sendo que hoje, os maiores ratos possuem no máximo 2 kg e são encontrados nas florestas da Nova Guiné e Filipinas.
Segundo estudos, a ilha do Timor Leste vem sendo habitada pelo ser humano há pelo menos 40 mil anos. Com a chegada do homem e o começo da agricultura na região, há aproximadamente 2 mil anos, é que teria se iniciado o processo de extinção do animal. Este teria se dado não só por alteração de seu habitat, mas provavelmente porque o rato gigante também era largamente utilizado na alimentação dos nativos.
Embora muitos não simpatizem com roedores, especialmente os ratos, esses animais são de incontestável importância para o ecossistema, sendo importantes dispersores de sementes e elementos fundamentais na manutenção de propriedades do solo.

Ossada do maior rato do Mundo (à esquerda) em comparação com o crânio de um tipo de rato atual.
Não só o curioso e bizarro Coryphomys, mas outros animais que hoje não passam de alguns centímetros um dia tiveram seus dias de glória sobre a Terra:
O estranho invertebrado artrópode Jaekelopterus rhenaniae, por exemplo, tratava-se de um tipo escorpião marinho – um euriptérido – que media cerca de 2,5 metros de comprimento e viveu há 390 milhões de anos. O tamanho de Jaekelopterus foi inferido a partir de sua garra de 46 cm encontrada na Alemanha. Para se ter uma idéia, os maiores escorpiões atuais chegam à cerca de 30 cm, mas em geral são muito menores do que isso. Os cientistas ainda acreditam que este europtérido tenha dado origem aos atuais escorpiões e aracnídeos. – Tenho que dizer que esse sutil e “simpático” animal me causa calafrios.

Imagem comparativa do tamanho de
Jaekelopterus rhenaniae e um homem
Outro fascinantes seres gigantes também já foram descobertos: aranhas, centopéias, libélulas e baratas de tamanhos descomunais; preguiças, cobras, crocodilos e aves tão grandes que inspirariam filmes de terror; porém muitos ainda estão a espera de serem desvendados. A ciência tem muito o que vasculhar e é incrível a possibilidade de reconstituir parte do fantástico cenário de vida de épocas e eras passadas. Reviver paleopesadelos!
Agora a pergunta que não quer calar: Por que esses animais atingiram esse grande porte? Bom, há muitas hipóteses. Para explicar o gigantismo dos artrópodes terrestres, por exemplo, há cientistas que afirmam que na época em que esses animais viveram, a atmosfera era abundante em oxigênio, o que teria favorecido parcialmente essa adaptação corporal. Já outros, acreditam que esse grande crescimento teria se dado pela falta de predadores como os vertebrados nos ambientes terrestres recém-colonizados. Para os vertebrados, no entanto, o gigantismo tem outras explicações, pode se dar por questões intrincadas de fisiologia, relação predador/presa e ainda há a interessante “regra de Cope”. Esse é um assunto que gera um intenso debate e que em outras oportunidades iremos dar continuidade aqui no Colecionadores.