

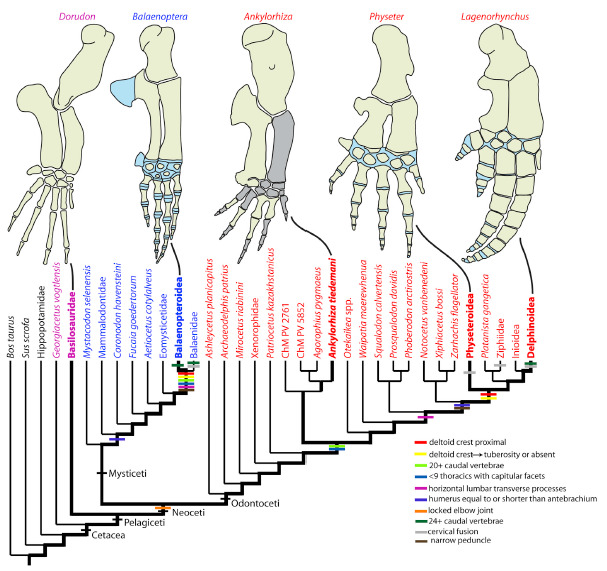
Nas últimas postagens você teve a oportunidade de conhecer alguns grandes cetáceos fósseis predadores, como Basilosaurus isis e Ankylorhiza tiedemani. Mas não só de caçadas sanguinolentas vivem esses animais. Bom, isso se você não for um cardume de lulas, peixes ou crustáceos...
Na postagem de hoje, vamos contar sobre como as baleias foram por um caminho um pouco diferente e se tornaram alguns dos maiores seres viventes do planeta, alimentando-se por filtração. Caso você seja parte do cardápio, é bom por sebo nas canelas, ou melhor, nas nadadeiras, porque a fome aqui é gigantesca.
Um balaio de baleia
Antes de qualquer coisa, é importante entender quem são as baleias filtradoras:
Em inglês, o termo whale (baleia) pode ser usado popularmente para se referir a qualquer espécie de cetáceo, seja este pequeno ou grande, com dentes ou não. Porém, também existe uma palavra para se referir especificamente às baleias filtradoras : baleen (algo como “barbas” ou “barbatanas”, em português).
“Baleen” refere-se particularmente aos Mysticeti ou “misticetos“, no bom português. Misticetos são os cetáceos dos grupos Balaenopteridae, Balaenidae, Eschrichtiidae, Neobalaeninae e alguns outros grupos fósseis. Estes grupos possuem barbatanas (ou barbas) no lugar dos dentes, que servem para filtração, e é por isso, que esses cetáceos são chamados de “baleias filtradoras” ou “baleias de barbatana” aqui no Brasil.
Também há nomes populares específicos para diferenciar certos grupos. Rorqual, por exemplo, refere-se somente às baleias da família Balaenopteridae, que incluem baleias-azuis, baleias-fin, baleias-sei, baleias-de-bryde, baleias-de-rice, baleias-minke e baleias-jubarte. Baleias verdadeiras, por sua vez, é o nome usado para denominar as Balaenidae, representas pelas baleias-francas-austrais, baleias-francas-do-atlântico-norte, baleias-francas-do-pacífico, e as baleias-da-groenlândia.
As outras duas famílias, Eschrichtiidae e Neobalaeninae, não ocorrem no Brasil, e normalmente são chamadas pelos nomes das espécies que as representam: baleia-cinzenta (Eschrichtiidae) e a rara baleia-franca-pigméia (Neobalaeninae).

Modos à mesa
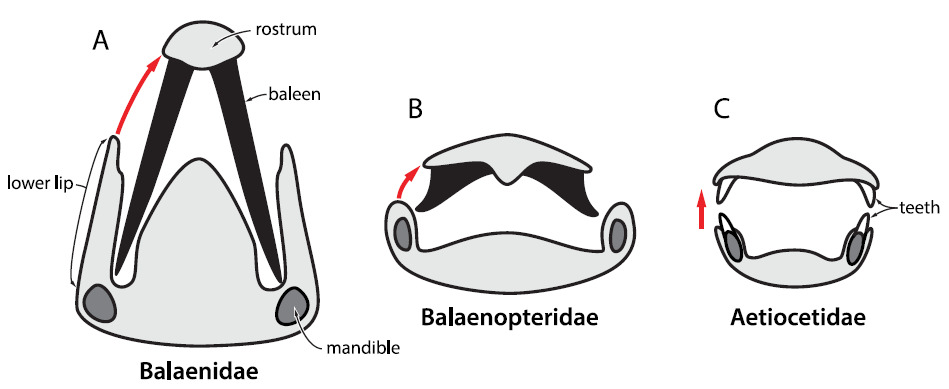
Cada grupo de baleia filtradora possui características anatômicas diferentes que refletem modos únicos de filtrar a água para se alimentar. Porém, pelo menos uma coisa é comum entre elas: as barbatanas, estruturas compridas e enfileiradas, feitas de queratina (mesmo material que forma nossos cabelos e unhas), presentes na boca desses animais. Tais barbatanas acabaram por substituir os dentes dos misticetos ao longo de sua evolução.

As barbatanas funcionam como uma escumadeira ou peneira, que permite que a água abocanhada durante a alimentação seja expulsa da boca, com a ajuda da língua, e que o alimento fique preso enquanto a água sai.
As rorquais possuem pregas na região ventral, que se expandem como o papo de um pelicano, permitindo com que elas possam abocanhar (engolfar) grandes quantidades de água com cardumes inteiros de pequenos peixes ou krill (um tipo de camarãozinho). Já as baleias-verdadeiras, são mais corpulentas, não possuem pregas ventrais e a sua boca é em forma de arco. Elas basicamente se alimentam filtrando a água enquanto nadam com a boca aberta.
As barbatanas das baleias-verdadeiras podem ter entre 2 metros de altura, até 5,2 metros de altura!
As baleias-cinzentas, por sua vez, abocanham a areia do fundo marinho filtrando-a em busca de crustáceos enterrados. Elas possuem sulcos na garganta em vez de pregas ventrais. As baleias-franca-pigméia, por fim, possuem uma mistura de características: boca em forma de arco como a das baleias verdadeiras, sulcos ventrais na garganta como as baleias-cinzentas e estilo de alimentação como as rorquais.

Mas sempre foi assim? Como será que as barbatanas surgiram? As formas de alimentação sempre foram as mesmas desde o começo da evolução dos misticetos? Como as espécies transicionais se alimentavam? Essas são perguntas que o registro fossilífero pode ajudar a responder…
Comendo com hashi, os “palitinhos japoneses“
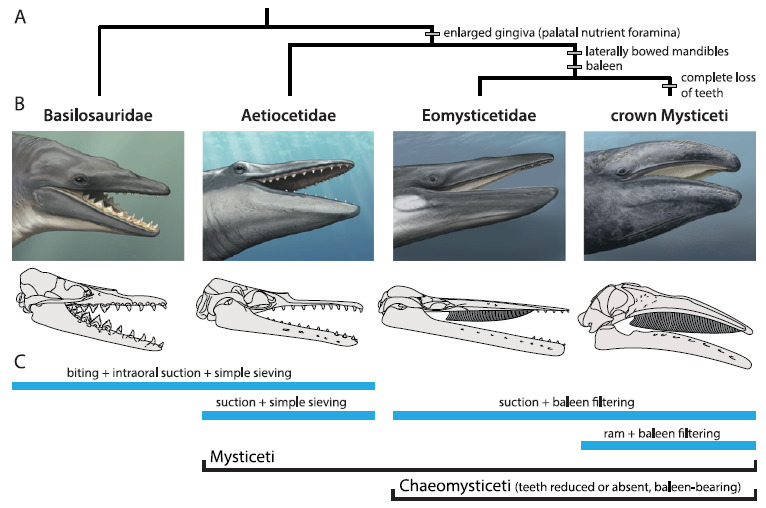
Os basilossaurídeos foram os primeiros cetáceos totalmente aquáticos. Eles eram grandes caçadores, inclusive de outros cetáceos, e tinham a boca cheia de dentes diferentões, como já citamos AQUI. A pergunta inevitável é: como essas “baleias primitivas” foram de caçadoras dentadas para as banguelas filtradoras que temos na atualidade? Bom, alguns achados fósseis das últimas décadas possibilitaram entender parte desse processo:
Em 2016, Felix G. Marx junto com outros colegas, analisaram estranhas ranhuras encontradas em dentes fósseis de um grupo de baleias “primitivas” chamadas de Aetiocetidae, comuns no Oligoceno.
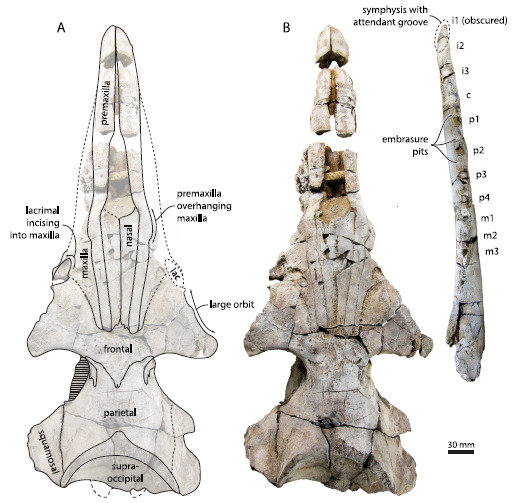
Eles analisaram um fóssil específico, encontrado em rochas da região de Washington, EUA. O espécime, datado do Oligoceno Superior demonstrou afinidade aos misticetos, mas possuía um padrão de desgaste nos dentes muito diferente, não compatível com a presença de barbatanas. Felix e colegas, depois de muitas análises, associaram os desgastes horizontais nos dentes a uma alimentação por sucção, sugerindo que as barbatanas dos misticetos teriam surgido mais tarde, próximo à origem das baleias filtradoras modernas. O que isso significa? Que antes de filtrar, os misticetos provavelmente teriam se alimentado sugando o alimento, como se faz quando comemos macarrão com hashi!
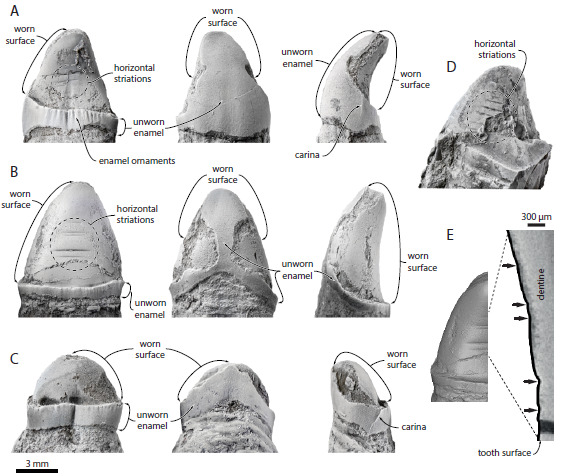
Figura 5. Padrões de desgaste em dentes, sugerindo alimentação por sucção em um aetiocetídeo (MARX et al., 2016).
As ranhuras horizontais observadas nos dentes foram comparadas com outros animais que também se alimentam por sucção na atualidade, como belugas, algumas baleias-bicudas, morsas e certas espécies de focas. Pelo padrão de desgaste, a língua deveria funcionar como um pistão, fazendo pressão para que a água e a(s) presa(s) fossem sugados rapidamente para dentro da boca, o que deveria causar estes desgastes.
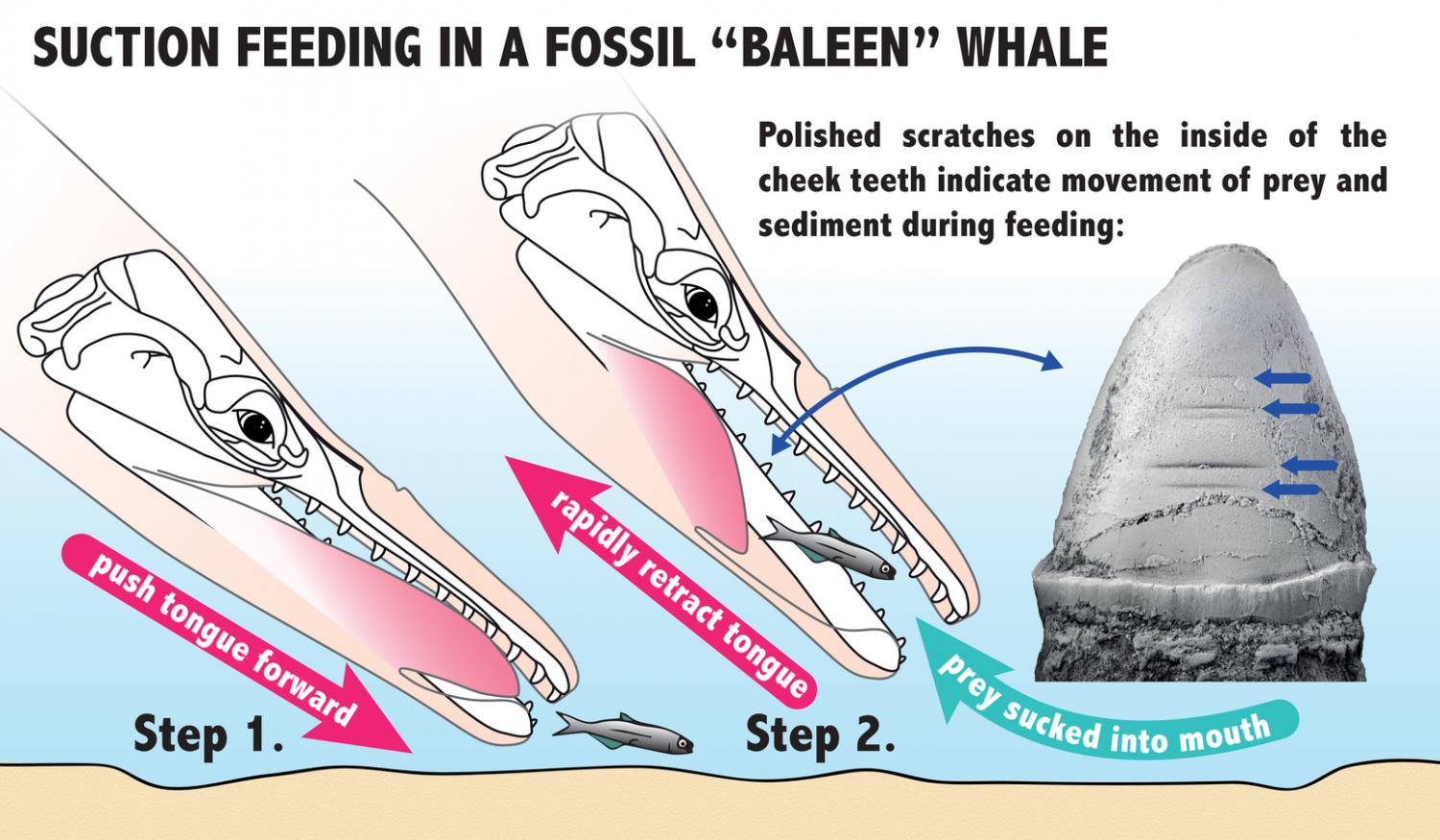
Também foram observadas marcas em alguns dentes, relacionadas com a expulsão da água e sedimentos que eventualmente eram sugados junto com o alimento. Os Aetiocetidae deveriam ficar com a boca meio aberta enquanto expulsavam tudo aquilo que não era alimento. Essas marcas são semelhantes às encontradas em certas espécies de mamíferos marinhos citados anteriormente, que também se alimentam por sucção.
Mas e agora? Onde as barbatanas aparecem nessa história, se os primeiros misticetos ainda tinham dentes e sugavam a comida, ao invés de morder tudo que passava pela frente? Mais uma vez, o registro fossilífero pode nos ajudar a entender essa história, mas agora vamos contar com a ajuda de análises em animais atuais para entender outra parte dessa saga evolutiva única.
Comendo com garfo e colher
Alguns autores sustentam que as barbatanas teriam aparecido em alguns misticetos extintos que ainda possuíam dentes. Eles descrevem que essa teria sido uma “fase de transição” evolutiva, em que esses cetáceos teriam exibido uma alimentação mista: de predadores ativo por sucção, que também seriam capazes de realizar filtragem. Achados fósseis do Oligoceno Superior, publicados em 2008, por Thomas A. Deméré e colegas, permitiram o reconhecimento de marcas no céu da boca de espécies fósseis de Aetiocetidae, que indicariam uma alimentação por filtração.

O material descrito por Thomas pertence a uma espécie denominada Aetiocetus weltoni. Nos fósseis foram encontradas ranhuras de inervação no palato (céu da boca) semelhantes àquelas observadas nas baleias filtradoras atuais. As inervações saem da região dos alvéolos dentários (local onde se encaixam os dentes) exatamente como nos misticetos atuais. Segundo os autores, isso seria uma evidência de que a espécie fóssil teria tido barbatanas.
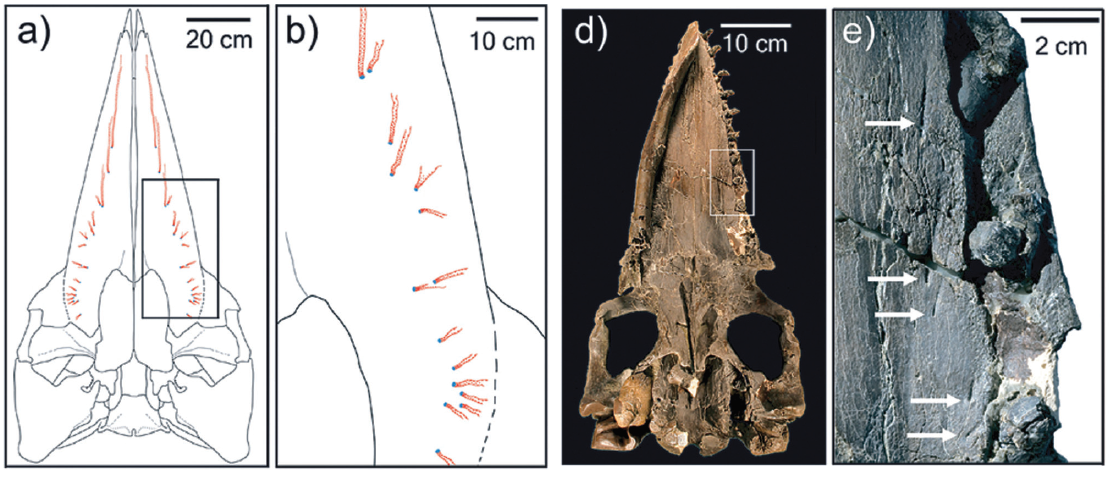
Inicialmente achava-se que as barbatanas poderiam ter surgido a partir de estruturas rígidas no céu da boca dos misticetos basais, chamadas de “cristas córneas palatinas”, também presentes nos artiodáctilos (cabras, bois, camelos, hipopótamos, etc.), grupo no qual os cetáceos são aparentados. Porém, as inervações encontradas nos fósseis observados por Thomas e colegas reforçaria outra hipótese.
Thomas e colegas observaram fetos de baleias filtradoras atuais e notaram que, apesar de “banguelas” quando adultas, a sua sequência de dentes se desenvolve em sua fase fetal. O seu crescimento é que é interrompido pela ação de alguns genes. Quando tais genes se ativam, os dentes são reabsorvidos pelo organismo, mantendo somente as inervações no palato geradas durante o seu crescimento inicial.

Durante a reabsorção dentária, queratina é secretada e é isso que forma as placas de barbatana no lugar dos dentes reabsorvidos. Essas placas crescem constantemente enquanto o filhote se desenvolve e, ao longo da vida, são desgastadas e desfiadas, ficando com a aparência que conhecemos.
A ordem dos talheres
Alguns autores, todavia, alertam: assim como a origem das penas em dinossauros não-avianos não marca a origem do vôo, o aparecimento dos canais de inervação no palaro dos misticetos não necessariamente indicaria a presença de barbatanas. Em vez disso, esses sulcos inervados em alguns aetiocetídeos fóssei poderiam, por exemplo, ter fornecido condições anatômicas e fisiológicas iniciais para o aparecimento posterior das barbatanas.
Felix G. Marx e colegas, em seu trabalho de 2016, apontam que a presença de dentes alternados, como os observados no fóssil descrito por Thomas e colaboradores em 2008, acabariam danificando as barbatanas, se essas estivessem presentes. Os mesmos autores também observam que a mandíbula das baleias filtradoras atuais é mais larga, o que permite com que as barbatanas não sejam danificadas durante o fechamento da boca do animal. Além disso, misticetos atuais também possuem uma adaptação muscular especial que permite com que a mandíbula rotacione levemente durante a oclusão, o que também dificulta o dano às barbatanas. Em aetiocetídeos, por sua vez, o tamanho da mandíbula e sua forma de abertura indicam que, qualquer barbatana, se presente, seria danificada durante o fechamento da boca.
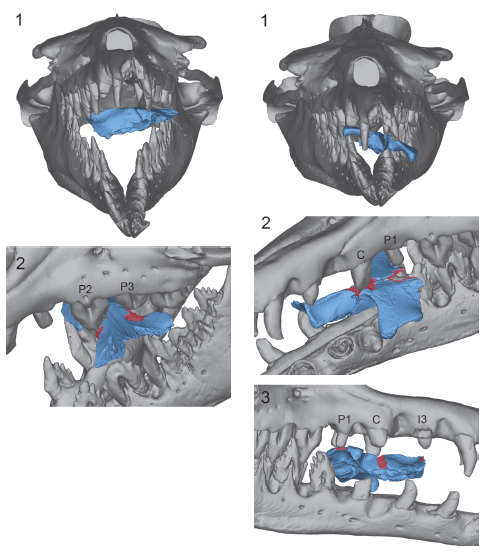
Felix e demais colegas até tentam dar uma chance ao modelo de barbatanas+dentes. Eles sugerem um cenário em que as barbatanas poderiam estar entre os dentes do animal e que, com o fechamento da boca, elas se dobrariam para dentro, similar ao que ocorre com as baleias-da-groenlândia. Todavia, ainda sim, a presença dos dentes alternados atrapalharia o dobramento e danificaria significativamente as estruturas. Os autores concluem, que as barbatanas não teriam surgido dessa forma. Talvez elas tenham surgido em outros grupos que tivessem uma mandíbula mais larga e/ou uma dentição reduzida ou inexistente.
Um jeito alternativo de comer
A capacidade de gerar sucção é fundamental para a maioria dos vertebrados aquáticos e é amplamente observada entre os mamíferos marinhos atuais. No entanto, até o trabalho de Felix e colegas, ela raramente havia sido associada à evolução dos misticetos. A sucção é muito útil na alimentação subaquática, pois facilita o transporte do alimento até o fundo da boca, onde ele será deglutido. Essa forma de adquirir alimentos provavelmente já estava presente em cetáceos basais, muito antes dos misticetos, mas esse é um comportamento relativamente difícil de se interpretar por meio de fósseis.
O uso de sucção na alimentação e provável ausência de barbatanas nos misticetos basais sugeriria um modelo diferente de evolução da alimentação por filtragem em baleias:
(1) Misticetos basais, incluindo aetiocetídeos, tinham tanto a dentição funcional, quanto a habilidade de usar sucção, herdada de cetáceos anteriores;
(2) A água ingerida por eles, como resultado da sucção, era expelida fundamentalmente com auxílio dos dentes;
(3) algum grupo de misticeto ancestral aprimorou suas capacidades de sucção ao longo do tempo, com o desenvolvimento de mandíbulas mais largas e tecidos moles associados (“calos” ou dobras na gengiva, lábios expandidos, etc.). O aperfeiçoamento desta capacidade teria favorecido a perda da dentição com o tempo;
(4) Com a perda da dentição, as barbatanas teriam evoluído e, gradualmente, sido selecionadas.
Este cenário seria mais plausível, pois exclui problemas potenciais de interferência entre uma dentição ativa e barbatanas. Ele explica também como os dentes poderiam ter sido perdidos, sem afetar o sucesso da alimentação. Além disso, ele está de acordo com a evidência observada no desenvolvimento de fetos de misticetos atuais, que mostra que o crescimento das barbatanas só se dá quando os dentes são reabsorvidos.
A sucção seguida de filtragem é uma forma de alimentação que pode limitar bastante o tamanho máximo da presa a ser capturada. O aperfeiçoamento dos tecidos moles associados à dentição, em especial a gengiva, é uma forma de lidar com este problema. Primeiro, um mecanismo para prensar a presa deve ter surgido nos misticetos basais e, depois disso, um aparato apropriado de filtragem – isto é, as barbatanas.
Uma adaptação observada nos botos-de-dall (Phocoenoides dalli), espécie vivente de cetáceo, pode ajudar a imaginar como o processo teria acontecido. Esses cetáceos odontocetos (não diretamente aparentados às baleias filtradoras) têm dentes rudimentares e possuem em sua boca, também, pequenos “dentes na gengiva”, semelhantes aos brotos iniciais das barbatanas dos misticetos. Essas estruturas ajudam os botos-de-dall a capturarem suas presas. Pode ser que essa adaptação seja uma pista de como se deu a perda gradual da dentição em misticetos e posterior surgimento das barbatanas.
A origem das barbatanas, adaptação chave das baleias filtradoras modernas (Mysticeti), marca uma transição profunda e única na história evolutiva dos vertebrados. Hoje, ela é um pouco melhor compreendida, porém, como vocês viram, ainda restam detalhes a serem desvendados sobre como esta belíssima crônica evolutiva se desenrolou. A compreensão da evolução dos “modos à mesa” das baleias pode estar, não só nos fósseis, mas em uma combinação entre estudos paleontológicos, genéticos e do desenvolvimento.
Referências:
CASTRO, Peter; HUBER, Michael E. Biologia marinha. AMGH Editora, 2012.
DEMÉRÉ, Thomas A.; MCGOWEN, Michael R.; BERTA, Annalisa; GATESY, John. Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Systematic biology, v. 57, n. 1, p. 15-37, 2008.
MARX, Felix G.; HOCKING, David P.; PARK, Travis; ZIEGLER, Tim; EVANS, Alistair R.; FITZGERALD, Erich M. G. Suction feeding preceded filtering in baleen whale evolution. Memoirs of Museum Victoria, v. 75, 2016.

















{kind=link}