Uma nova e interessante proposta para explicar as relações evolutivas dos dinossauros e principalmente, a origem dos dinossauros ornitísquios, grupo que inclui o Triceratops e os famosos dinossauros “bico de pato”, acaba de ser apresentada por dois pesquisadores brasileiros da Universidade Federal de Santa Maria (UFSM), RS, Rodrigo Temp Müller e Maurício Garcia. O estudo foi publicado na revista científica Biology Letters e agita ainda mais a discussão sobre as relações evolutivas dos dinossauros.
Arte de Márcio Castro
Não faz muito tempo que um estudo liderado por um pesquisador britânico chamado Matthew Baron, sacudiu a comunidade paleontológica e abriu espaço para novas discussões sobre uma questão fundamental no estudo de dinossauros: a relação evolutiva entre os grandes grupos desses animais.
Em 2017, Baron e colaboradores ressuscitaram uma antiga hipótese sobre a relação dos dinossauros, que propunha que o grupo que inclui os dinossauros carnívoros, como o Tyrannosaurus, era mais proximamente relacionado ao grupo dos dinossauros ornitísquios, aquele que reune Triceratops, Stegosaurus e os chamados “dinos bico de pato”. A hipótese de Baron e colegas veio com algumas novidades, mas já havia sido considerada no passado, tendo sido descartada por diversos estudos em detrimento da proposta mais clássica, que reúne dinossauros carnívoros e os grandes dinossauros “pescoçudos ” em um mesmo grupo chamado Saurischia.
O agito causado por esse artigo foi tanto, que, na época, muito se falou na mídia popular, inclusive, como se tudo que sabíamos sobre dinossauros tivesse que ser reescrito. Pouco tempo passou, todavia, até a publicação de um outro estudo, rebatendo a proposta de Baron e colaboradores. Estudo esse, liderado por um pesquisador brasileiro, da USP de Ribeirão Preto, Prof. Max Langer. O estudo liderado por Langer apontava inconsistências no trabalho de Baron e concluía que a hipótese clássica da divisão dos dinossauros ainda se sustentava com prioridade, apesar de a proposta de Baron não poder ser totalmente descartada, já que era somente “um pouco menos provável que a hipótese tradicional”. Leia mais sobre isso aqui.
A história é longa e a disputa continuou em alguns estudos subsequentes, mas aonde eu quero chegar é que, toda essa discussão ressaltou novamente uma verdade muito incômoda para os estudiosos de dinossauros: nós conhecemos muito pouco sobre os primeiros ornitísquios e não conseguimos dizer com certeza como eles se relacionam com os outros dinossauros. E é aí, finalmente, que o trabalho recém-publicado pelos colegas da UFSM pode ajudar.
Os “caçadores de dinossauros” da UFSM
Rodrigo Temp Müller e Maurício Garcia são dois pesquisadores muito privilegiados, pois têm a honra de estar justamente no local que provavelmente foi o berço dos dinossauros, há aproximadamente 230 milhões de anos. Os fósseis de dinossauros mais antigos conhecidos no mundo são encontrados em rochas dessa idade na Argentina, no sul do Brasil e na porção sul do continente africano. Justamente por estarem trabalhando sobre essas rochas, esses pesquisadores têm acesso direto a fósseis fantástico, que contam a história da aurora desse grupo tão fascinante de animais.
Rodrigo Temp Müller e Maurício Garcia
Atualmente, Rodrigo T. Müller e Maurício Garcia, junto a outros pesquisadores, trabalham vinculados ao Centro de Apoio à Pesquisa Paleontológica (CAPPA-UFSM), em São João do Polêsine, RS, onde se dedicam a estudar a história dos primeiros dinossauros.
A nova hipótese de Müller e Garcia
Verdade seja dita, aorigem dos dinossauros ornitísquios é um grande mistério. Enquanto os fósseis mais antigos de dinossauros saurísquios datam de estratos do meio do Período Triássico, entre 245 e 230 milhões de anos, os fósseis mais antigos de ornitísquios são um pouco mais recentes, da transição Triássico-Jurássico, e têm aproximadamente 200 milhões de anos. Explicar esta ocorrência usando a hipótese tradicional sobre da relação dos dinossauros é um pouco constrangedor, pois implica na existência de “linhagens fantasmas” de ornitísquios, que teriam existido entre ‘pouco tempo depois da origem dos dinossauros’, por volta de 240-230 milhões de anos, até cerca de 206 milhões de anos atrás, idade do fóssil mais antigo de ornitísquio conhecido. Resumindo: ornitísquios necessariamente teriam que ter existido entre ~240 e 206 milhões de anos, mas não temos evidências dos fósseis deles.
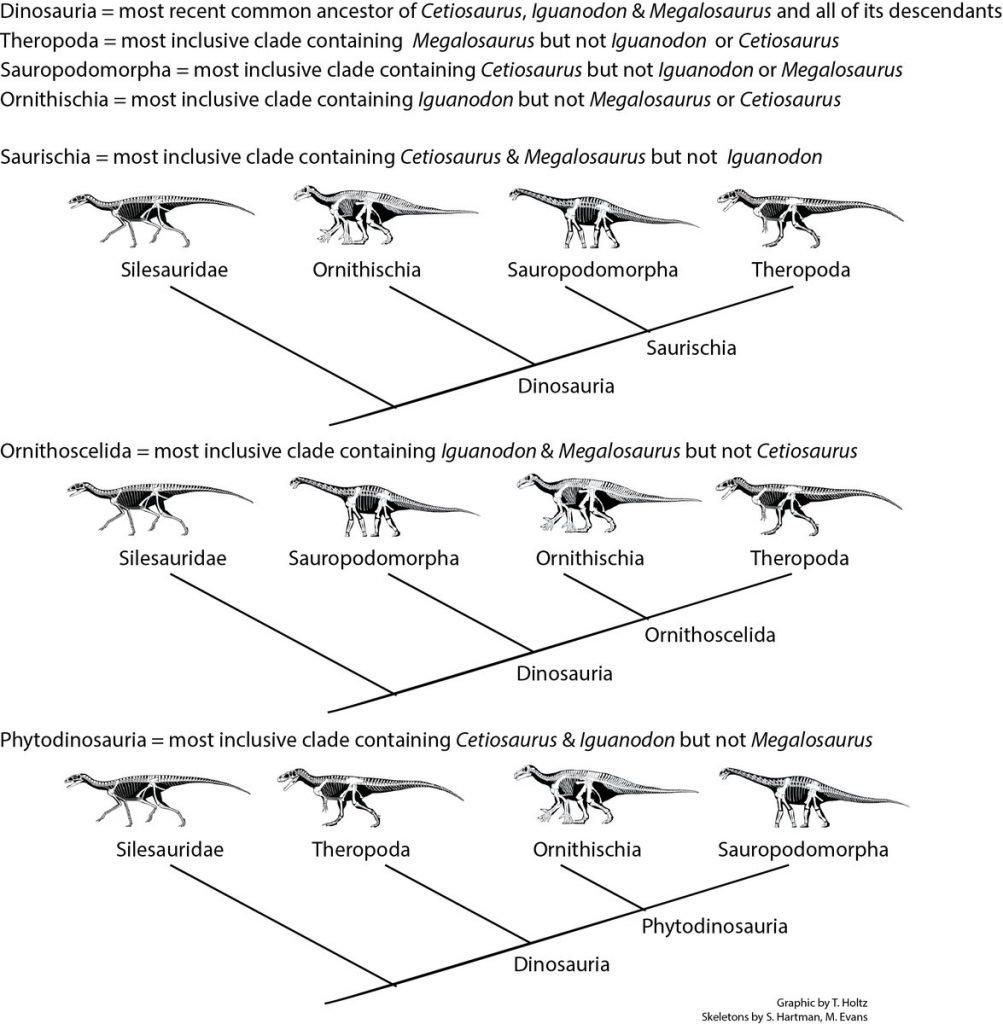
A nova hipótese proposta por Müller e Garcia solucionaria esse problema de maneira muito elegante, com algo que sempre esteve bem debaixo do nariz dos pesquisadores: os “Silesauridae”.
As criaturas conhecidas como ‘silessaurídeos’ compõe um grupo de organismos extintos normalmente considerados como parentes muito próximos dos dinossauros. De fato, eles e os primeiros dinossauros são extremamente parecidos, tanto em forma, hábitos, quanto em tamanho (imagem).
Fósseis de ‘silessaurídeos’ são encontrados no Brasil, como é o caso de Sacisaurus agudoensis, um pequeno animal, com cerca de 1,5m de comprimento, que viveu há 225 milhões de anos onde hoje é o Rio Grande do Sul.
Reconstituição artística de Sacisaurus agudoensis por Rodolfo Nogueira.
O que Müller e Garcia fizeram em seu estudo foi testar as relações evolutivas dos grandes grupos de dinossauros, incluindo uma ampla amostragem de silessaurídeos (até então tidos como grupo irmão de Dinosauria) e diversas espécies de dinossauros basais. Eles fizeram uma análise filogenética abrangente, que juntou, pela primeira, vez os dados de espécies há tempos conhecidas, com as espécies mais recentemente descritas. Os resultados encontrados por eles foram surpreendentes…
O que uma análise filogenética faz, basicamente, é testar as relações evolutivas dos organismos por meio da comparação de uma extensa matriz de dados sobre eles. Estes dados podem ser genéticos ou morfológicos, ou os dois, por exemplo. No caso dos fósseis, os pesquisadores normalmente usam dados morfológicos (da forma) para realizá-las, pois informações genéticas só ficam preservadas em materiais relativamente recentes (de até algumas centenas de milhares de anos). O produto de uma análise filogenética são um conjunto de “árvores evolutivas” possíveis, mostrando todas as relações prováveis entre os organismos analisados. Algumas árvores são estatisticamente mais plausíveis que outras e, normalmente são essas as consideradas mais atentamente pelos pesquisadores.
Uma incrível descoberta
O que Müller e Garcia recuperaram em suas análises foi algo diferente de tudo antes proposto e muito excitante para os estudiosos de dinossauros, já que tem o potencial de explicar várias questões sobre o início da história evolutiva do grupo. Em especial, o tal mistério das “linhagens fantasmas” de ornitísquios.
De acordo com os resultados do estudo, os chamados ‘silessaurídeos’ seriam, na verdade, uma série de espécies ordenadas sucessivamente na base de Ornithischia. Ou seja, linhagens que teriam gradualmente acumulado características típicas de ornitísquios ao longo de milhões de anos. Essa hipótese preencheria o intervalo evolutivo antes “assombrado” pela ideia de “linhagens fantasmas”.
Os “silessaurídeos” seriam os dinossauros ornitísquios basais “que faltavam”! De acordo com a hipótese recuperada na análise de Müller e Garcia (2020), ‘Silesauridae’ seriam um grupo parafilético, na base de Ornithiscia. Sob esta perspectiva, Sacisaurus voltaria a ser um dinossauro.
Imagem do artigo de Müller e Garcia (2020).
Outros autores já haviam apontado uma possível relação entre silessaurídeos e ornitísquios, mas o cenário evolutivo encontrado por Müller e Garcia é inédito. Os ornitísquios clássicos, segundo o novo estudo, teriam evoluído a partir de formas típicas de silessaurídeos por meio de mudanças graduais ao longo do tempo, partindo inclusive, de uma possível forma carnívora. O que assemelharia o cenário evolutivo dos ornitísquios ao dos saurísquios sauropodomorfos (veja este vídeo aqui para entender).
Todos os ornitísquios conhecidos até o momento apresentam características que os vinculam a uma dieta herbívora, todavia os mais antigos ‘silessaurídeos’ tinham dentes pontiagudos e afiados, possivelmente adaptados para uma dieta carnívora.
Os pesquisadores responsáveis pela pesquisa reconhecem que ainda é cedo para que a questão relacionada a origem dos dinossauros ornitísquios possa ser considerada totalmente respondida. A nova hipótese de Müller e Garcia agora deverá continuar sendo testada à medida que novos fósseis foram descobertos e novas interpretações realizadas.
Nós aqui também estamos de olho.
A nova proposta de Müller e Garcia (2020). Arte de Márcio Castro.Como ficaria a configuração da nova árvore evolutiva dos dinossauros de acordo com o estudo de Müller e Garcia (2020). Arte de Márcio Castro.
Se você se interessou pelo estudo, você pode ler ele na íntegra AQUI.
Leia também a postagem no blog do parceiro ‘Coelho Pré-Cambriano’: AQUI.
Referência:
Müller, R.T. & Garcia, M. 2020. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biology Letters, https://doi.org/10.1098/rsbl.2020.0417
Você já se perguntou “como era a cara dos nossos ancestrais, antes deles serem o que somos”? Por exemplo, que cara teria o primeiro hominídeo? Ou o primeiro primata?
Essa pergunta habita o nosso imaginário, principalmente quando diz respeito aos nosso ancestrais e, na maioria das vezes, quem pode nos ajudar a obter essas respostas são os pesquisadores que trabalham com o passado, como os paleontólogos.
Onde sua imaginação te levaria se eu te perguntasse: que cara tinha o primeiro mamífero? Muitos talvez tenham pensado nos grandes mamíferos do passado, como os mastodontes (como Stegomastodon waringi), ou os poderosos tigres-dentes-de-sabre (como Smilodon), ou ainda nas preguiças enormes (como Eremotheriumlaurillardi) e tatus gigantes (como Glyptodonclavipes). Porém, sinto lhe informar, que você viajou pouco no tempo.
Uma preguiça gigante (Scelidodon sp.) e um tatu gigante (Doedicurus sp.), ambos encontrados na América do Sul em rochas datadas do Pleistoceno, entre 2,5 milhões e 11,7 mil anos atrás. Artes de Jorge Blanco (Forasiepi, Martinelli, 2007).
Quando pensamos em um mamífero, o grande grupo de animais ao qual nós, os seres humanos, pertencemos, fica difícil escolher um modelo que represente o todo. Vemos hoje em dia, a enorme diversidade do grupo, que foi capaz de ocupar praticamente todos os ambientes do nosso planeta, das savanas quentes do Brasil e da África, às geleiras mais frias do pólo-norte, das montanhas mais altas do Himalaia, às profundezas do oceano, dos céus, ao interior de cavernas e do solo. Em todos esses ambientes você encontra um exemplo diferente de mamífero. Este grupo de animais se diversificou de tal forma e foi tão moldado pelos ambientes que colonizaram, que é difícil considerar que um elefante, um morcego e um golfinho pertençam ao mesmo grupo e sejam parentes. Talvez, isso se deva ao fato de que a diversidade de formas dos mamíferos hoje é maior em relação aos outros grupos de tetrápodes viventes. Pense nas aves ou nos lagartos ou nos crocodilos, que apresentam, na atualidade, uma variedade bem menor de formas e tamanhos do que os mamíferos (no passado não foi assim, mas esta é outra história). Pensando em tudo isso, qual animal você escolheria para representar os mamíferos? Que mamífero vivo hoje você diria que se assemelha mais ao ancestral de todos os mamíferos, ao primeiro mamífero?
Temos certeza que sua imaginação te deu várias opções, mas, sem querer te decepcionar, a cara do primeiro mamífero seria mais parecida com a de um musaranho ou de uma cuíca (não, não estamos falando do instrumento! Estamos falando do marsupial… Colocamos uma foto abaixo pra ajudar).
Filhote de cuíca (Didelphimorphia) – Foto dos autores.
O primeiro mamífero era um bicho pequeno, mais ou menos do tamanho de um pequeno gambá, correndo por entre as folhagens de uma floresta, durante uma noite quente do Jurássico (sim, a história dos mamíferos começa no Jurássico).
Atualmente, por consenso, o táxon apontado como o ‘primeiro mamífero’ é Morganucodon, um organismo fóssil encontrado nos EUA, Europa e China. Queremos chamar a atenção aqui para a expressão “atualmente apontado”, porque estes consensos taxonômicos podem mudar a luz de novos estudos, fósseis e evidências.
Reconstrução artísitica de Morganucodon. Seus fósseis são encontrados principalmente em Wales (Reino Unido) e na China, além de outras partes da Europa e América do Norte, em afloramentos Jurássicos. Imagem de FunkMonk (Michael B. H.).
O grupo chamado de ‘Mammalia’ (ou “mamíferos”, em bom português), é definido por um conjunto de características morfológicas compartilhadas por todos os seus membros. Colocando de forma mais simples: pra você ser um mamífero, você tem que ter, ou ter tido, um conjunto de características físicas apontadas como “coisa de mamíferos”. Mas tem um problema aqui. Vários organismos fósseis, muito próximos dos mamíferos já tinham algumas dessas “características típicas de mamíferos”. Isso é um pesadelo para muitos pesquisadores, que acabam por discutir e rediscutir definições…
A definição mais atual e com maior consenso, é a definição filogenética de mamífero, que englobaria Morganucodon e todas as espécies viventes de mamíferos (placentários, marsupiais e monotremados). Nessa definição, varias espécies de mamíferos extintos, que viveram durante a era Mesozoica, estão inclusas no grupo. Basicamente, isso significa que todos os animais que são agrupados numa árvore filogenética entre Morganucodon e os mamíferos atuais, são considerados mamíferos (calma, calma, a gente coloca uma figura, só olhar aí embaixo). Mas, essa definição também é bastante discutida, principalmente porque Morganucodon foi “eleito” como o primeiro mamífero, ou seja, essa é uma escolha arbitrária. Essa problemática de “eleger um primeiro” não é exclusiva dos mamíferos, esse é um conflito constante nos estudos sistemáticos e evolutivos, já que as formas biológicas formam um contínuo, quem tenta classificá-las em grupos artificiais somos nós.
No fim, cada novo achado acrescenta uma nova peça a esse quebra cabeça da evolução e as definições se atualizam com o tempo.
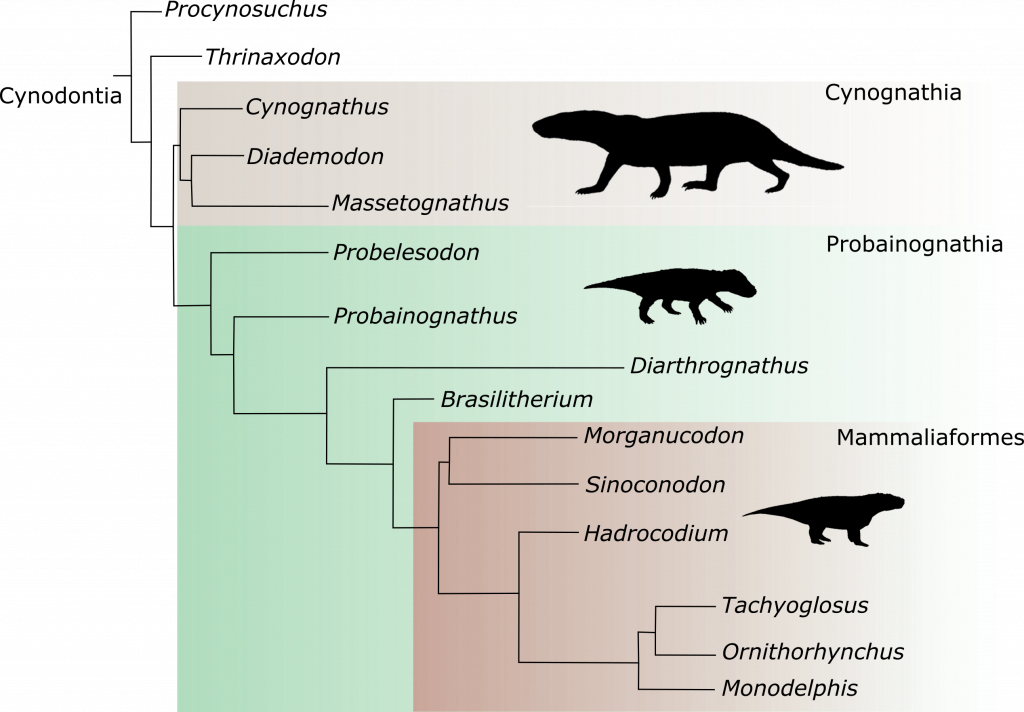
Filogenia simplificada dos cinodontes. Aqui estão apenas algumas poucas espécies da grande diversidade de cinodontes. Note que o grupo que Morganucodon é considerado o início do grupo dos mamíferos, portanto, todos que vierem depois deste grupo na árvore filogenética são considerados mamíferos. E um destaque para Brasilitherium, um fóssil brasileiro que é hoje tido como o fóssil mais relacionado ao grupo dos mamíferos. Modificado de Lautenschlager et al. 2016.
Quais são características presentes hoje nos mamíferos que definem o grupo como tal? Certamente você já ouviu que são as glândulas mamárias, os três ossículos do ouvido, entre outras. Mas para saber mais sobre elas, precisamos voltar no tempo. Mais precisamente, até os períodos Permiano e Triássico (entre cerca de 298 a 201 milhões de anos atrás), quando tais “características de mamífero” começam a ser observadas, gradualmente, em formas mais basais de animais aparentados dos mamíferos.
Durante a transição entre o Permiano e o Triássico, a Terra passou pelo seu maior evento de extinção, conhecido como a Extinção Permo-Triássica. Este evento foi bem maior do que a famosa extinção que dizimou os dinossauros. Essa tal Extinção Permo-Triássica foi tão grande, que causou um “reset” na fauna e na flora do planeta. Durante o final do Permiano (cerca de 255 milhões de anos atrás), os primeiros fósseis de criaturas conhecidas como cinodontes são registrados. Porém, é durante o Triássico que esses animais começam a brilhar no cenário biológico. Infelizmente, todos os holofotes acabam por se voltar para os dinossauros no final deste período, mas, o mundo dá voltas, como vocês verão.
Os cinodontes apresentavam uma grande diversidade de formas e tamanhos durante o Triássico e alguns já apresentavam algumas das tais “características mamalianas”. O curioso é que essas características não estavam presentes somente na linhagem que deu origem aos mamíferos. Alguns grupos de cinodontes completamente extintos, de uma linhagem paralela a nossa (mammaliana), também apresentavam algumas dessas características, que hoje, são consideradas como “coisa de mamífero”. Essa é a razão pela qual o debate sobre a origem dos mamíferos está sempre se modificando atualmente… Uma vez que vários grupos paralelos apresentam características mamalianas, é difícil associar com segurança, que determinado grupo de cinodontes deu origem aos mamíferos ou não.
Voltando para o assunto “que cara teria o primeiro mamífero?”, você deve estar se perguntando agora “que cara teriam os cinodontes?”. Se você pensou no musaranho ali em cima… você não está de todo errado, porém se você prestou atenção neste texto, você já sacou que eles têm uma grande diversidade de formas, e pasmem, em termos fósseis, o Brasil é um dos países que apresentam a maior diversidade de cinodontes do mundo! Todos eles provenientes do Rio Grande do Sul, o local que apresenta as formações de idade triássica mais fossilíferas do país. O Brasil trouxe ao mundo, por exemplo, os Brasilodontideos, o atual grupo apontado como o clado de origem dos mamíferos.
A Diversidade de Cinodontes Brasileiros
Antes de tudo, a gente precisa entender como são separados os cinodontes. Basicamente, existem dois grandes grupos dentro do grande grupo Cynodontia, os Cynognathia e os Probainognathia. Calma, a gente vai explicar um pouquinho de cada grupo abaixo:
Cynognathia inclui organismos completamente extintos. Eles eram em sua maioria herbívoros/onívoros, com exceção de apenas uma espécie, que era carnívora. Eram bichos relativamente grandes, variando do tamanho de um cachorro pequeno até o maior de todos, que podia ter mais de 2 metros de comprimento e pesar cerca de 200kg. Neste grupo existem organismos que já apresentavam algumas características que podem ser interpretadas como “coisa de mamífero”, por exemplo, uma das principais características do grupo (e que pode ser comparável a mamíferos), é a enorme complexidade dos dentes pós-caninos. Os mamíferos possuem um padrão dentário altamente especializado, chamado de tribosfênico. Os Cynognathia, embora não tivessem padrão tribosfênico, possuíam especializações dentária até então não encontradas em outros grupos de Synapsidas. Além da grande especialização dos dentes, recentemente foi encontrado em um cinodonte Cynognathia, chamado de Menadon, com um padrão de dente hipsodonte, de crescimento contínuo, tipo os encontrado hoje em mamíferos como o cavalo e roedores (se você não sabia disso, aqui vai mais uma curiosidade, o dente do seu ratinho cresce pra sempre…por isso ele está sempre roendo algo. Não só ele, como vários outros animais). Essa ocorrência de dente hipsodonte no Menadon é única, e este é o único gênero além dos mamíferos com esse padrão de dente. O mais interessante, é que o grupo de Menadon foi completamente extinto, então a característica que era tida como exclusiva de mamíferos, já tinha aparecido na história dos cinodontes muito tempo antes! Infelizmente, toda a linhagem de Cynognathia foi extinta, então nunca teremos a oportunidade de ver um vivo e verificar como eles realmente seriam.
Cynognathia, os fósseis desse grupo são muito abundantes na Argentina, como o Massetognathus pascuali, e no Brasil, onde encontramos várias espécies em abundância, como Menadon e Santacruzodon. Abaixo a reconstrução de duas espécies de Cynognathia em um típico ambiente do Triássico, com destaque pra aparência que já lembraria muito a de um mamífero atual. Imagens: Massetognathus (foto do autor) Menadon (Melo et al. 2019) e a reconstrução artística por Voltaire D. P. Neto.
O segundo grupo, Probainognathia, abrange uma variedade de formas gigantesca, já que Mammalia está inclusa neste grupo. Mas, levando apenas os fósseis em consideração, o grupo apresentava mesmo assim uma diversidade de tamanho e de hábitos enorme, variando de um bicho com o tamanho de um cachorro grande (como o Aleodon, que podia ter mais de 1,5 metros), até os Brasilodontídeos (que tinham o tamanho de um pequeno gambá, com cerca de 15cm). Os animais desse grupo são, em sua maioria, classificados como insetívoros ou seja, eles comiam insetos, porém, alguns pesquisadores apontam que eles poderiam ser oportunistas (onívoros, assim como os gambás atualmente), com alguns exclusivamente carnívoros, como o Trucidocynodon. Neste grupo estão incluídos os Brasilodontidae, atualmente tido como grupo irmão de mamíferos, mas que pode ter sido o grupo de cinodontes que deu origem a nós, os mamíferos.
Probainognathia. Artes de Jorge Blanco (Martinelli et al. 2016; Guignard et al. 2019).
A parte mais fantástica disso tudo, é que muitos desses bichos faziam parte da fauna triássica do Brasil. Eles estão entre os achados fósseis do Rio Grande do Sul, onde é encontrada a maior diversidade de Cynognathia do mundo, além de alguns dos registros mais importantes de Probainognathia, como os já mencionados Brasilodontideos. Talvez, devido ao pequeno tamanho, os cinodontes acabem por perder espaço para os grandes dinossauros na mídia e também no imaginário das pessoas… Apesar disso, imaginar um “pequeno musaranho”, correndo de um dinossauro, numa noite quente do Triássico, está carregado de informações sobre como nós, os mamíferos, conseguimos nos tornar o que somos hoje. Enfim, agora você sabe como era “a cara dos primeiros mamíferos” e também como os fósseis do Brasil são importantes para contar essa história.
Referências
Forasiepi A, Martinelli A. Bestiario fósil: mamíferos del pleistoceno de la Argentina. Albatros; 2007.
Guignard ML, Martinelli AG, Soares MB. The postcranial anatomy of Brasilodon quadrangularis and the acquisition of mammaliaform traits among non-mammaliaform cynodonts. PloS one. 2019 May 10;14(5):e0216672.
Lautenschlager S, Gill PG, Luo ZX, Fagan MJ, Rayfield EJ. The role of miniaturization in the evolution of the mammalian jaw and middle ear. Nature. 2018 Sep;561(7724):533-7.
Martinelli AG, Soares MB, Schwanke C. Two new cynodonts (Therapsida) from the Middle-Early Late Triassic of Brazil and comments on South American probainognathians. PloS one. 2016 Oct 5;11(10):e0162945.
Melo TP, Ribeiro AM, Martinelli AG, Soares MB. Early evidence of molariform hypsodonty in a Triassic stem-mammal. Nature communications. 2019 Jun 28;10(1):1-8.
Olá caros leitores, depois de um breve período de pausa nas postagens, voltamos com grande estilo. Hoje apresento a vocês um interessante texto redigido pelo Mestrando em Zoologia do Museu Nacional/UFRJ Geovane Alves de Souza, O assunto abordado se refere as primeiras discussões históricas sobre as hipóteses filogenéticas dos Amniotas (grupo que tradicionalmente inclui os répteis, aves e mamíferos e suas formas relacionadas) e a importância dos fósseis para um melhor entendimento da evolução das espécies. Então, sem mais delongas, vamos ao texto!
Editado em 19/08/2018.
Em plena Era da Filogenômica, na qual o DNA possui papel central na busca pela compreensão da evolução da vida na Terra, é comum pensarmos nos fósseis como uma fonte de dados um tanto quanto ultrapassada, trabalhosa e que demanda muito tempo para estudar. A diretora do Jurassic World, Claire Dearing (encenada pela atriz Bryce Dallas) enfatiza isso muito bem em sua fala: “Aprendemos mais com a genética em 10 anos do que em um século, escavando”. Hoje é consenso no meio acadêmico a importância dos vestígios da vida pretérita, os fósseis (para melhores detalhes acerca de sua definição veja aqui), quando inferimos relações de parentesco dos organismos viventes. Contudo, nem sempre os fósseis tiveram sua importância reconhecida e passando por momentos de glória e queda ao longo dos últimos séculos.
Após a publicação da obra A Origem das Espécies por Meio da Seleção Natural por Charles Darwin em 1859, no qual o autor defendia as teorias de Evolução Biológica, Seleção Natural e Ancestralidade Comum, os fósseis passaram a desempenhar um papel chave na compreensão de como a vida evoluiu. Contudo, foi na metade do século XX, que o registro fossilífero enfrentou uma queda brusca de sua supremacia. O responsável foi o advento de uma nova maneira de se estudar a evolução: a Sistemática Filogenética de Willi Hennig (1950). A nova metodologia e filosofia da sistemática nos seus primeiros anos de existência não exigia a necessidade de determinar uma dada espécie fóssil conhecida como ancestral entre duas linhagens. Podíamos estudar a evolução das espécies viventes, tratando o ancestral comum entre elas como uma espécie hipotética. Esse modo de vislumbrar as árvores filogenéticas (diagramas ramificados que representam a evolução de uma linhagem e que são gerados a partir de uma análise computacional ou análise filogenética), conhecido como Modelo Cladogenético, é antagônico ao antigo modelo vigente, o Anagenético. De fato, determinar em qual ponto exato da evolução de uma linhagem uma espécie fóssil esta inserida não é uma tarefa fácil, eu diria que um tanto quanto impossível, a menos que tenhamos uma máquina do tempo para voltarmos e acompanharmos o passo a passo da evolução de determinada linhagem ao longo dos milhares de anos. Muitas críticas contra o uso dos fósseis foram levantadas na época. O próprio Hennig reconhecia que os dados fósseis poderiam ser úteis na hora de conduzir uma análise filogenética. Contudo, devido à tamanha incompletude do registro fossilífero, ou seja, tanta informação biológica era perdida no processo de formação de um fóssil, que estes deveriam ser preferivelmente menos utilizados na hora de reconstruir as relações de parentesco.
Petterson (1981) mostrou, através de vários exemplos, o quanto os fósseis prejudicavam a compreensão sobre as hipóteses de evolução dos animais. Ax em 1987 defendeu em seu livro The Phylogenetic System que os dados fósseis são tão incompletos que as árvores deveriam ser construídas com base apenas nos grupos viventes e só depois que a análise computacional fosse feita é que se deveriam adicionar os fósseis. Dessa maneira e com muito sucesso, os críticos rapidamente conseguiram marginalizar o uso dos dados paleontológicos nos estudos da evolução das linhagens de organismos viventes.
A Hipótese Clássica da evolução dos Amniotas
Inúmeras árvores foram construídas ao longo dos anos seguintes, a maioria delas ignorando as informações provindas dos fósseis. O estudo que mais me chamou a atenção foi o trabalho clássico de Gardiner em 1982, no qual este autor tentou reconstruir a até então, pouco compreendida história evolutiva dos amniotas. Amniota é um grupo de animais vertebrados que possuem, dentre muitas características, uma membrana extraembrionária ao redor do feto chamada de amnion, membrana a qual é fundamental para a independência da água do ambiente durante o desenvolvimento do filhote no ovo, permitindo que estes animais colonizassem completamente o habitat terrestre. Estamos falando então da maioria esmagadora de vertebrados terrestres (e os que secundariamente retornaram ao ambiente aquático) que dominaram a Terra: desde as formas extintas famosas como dinossauros, pterossauros, ictiossauros, plesiossauros até as espécies contemporâneas de tartarugas, crocodilos, lagartos, serpentes, aves e mamíferos.
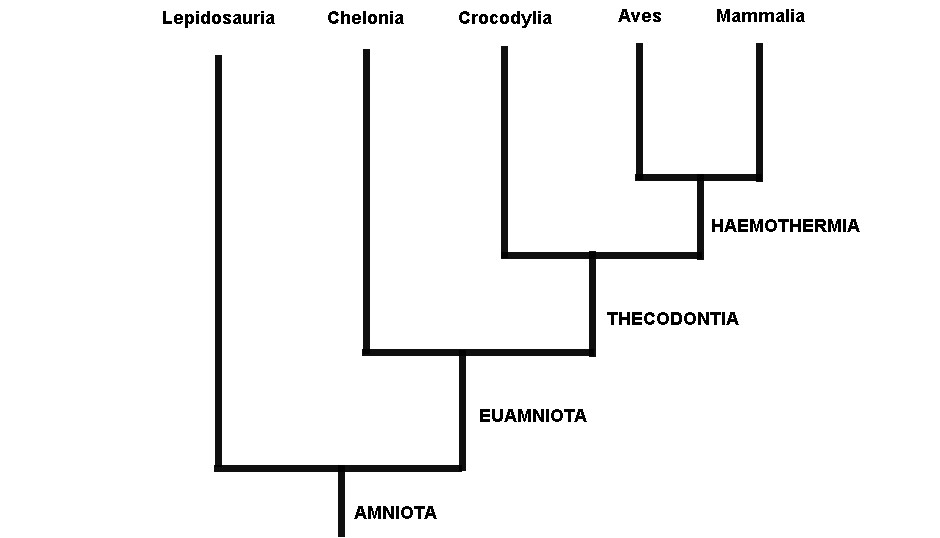
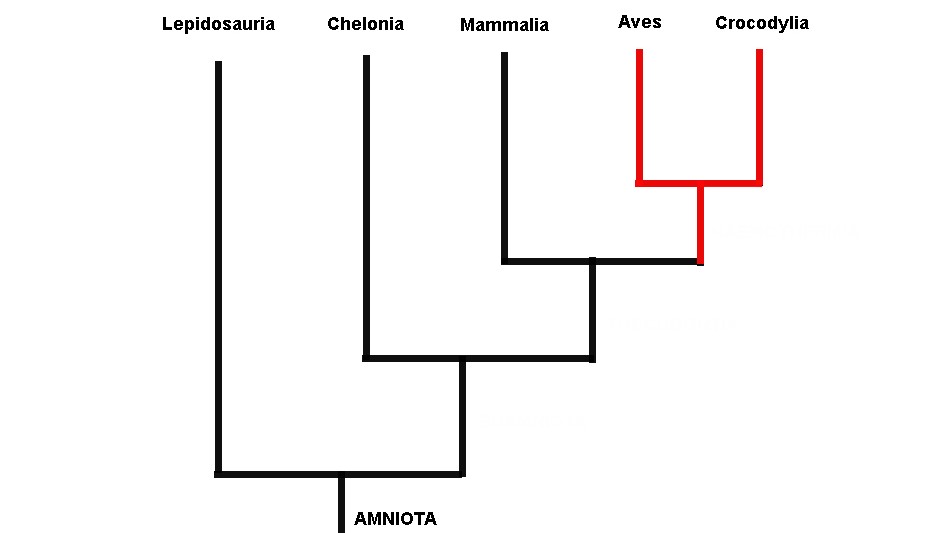
Gardiner utilizou vários dados morfológicos dos cinco grupos de amniotas viventes em suas análises (tartarugas, lagartos, jacarés, mamíferos e aves). Gardiner observou que as aves e os mamíferos eram evolutivamente relacionados, sendo agrupados por uma série de características que eram adaptações às suas altas taxas metabólicas (metabolismo alto leva a uma temperatura corpórea alta, sendo estes animais equivocadamente chamados de animais de “sangue quente”). Gardiner ressuscitou o antigo termo Haeomothermia para nomear o grupo de animais de “sangue quente” formado por aves e mamíferos (o termo vem de homeotermia, do grego homo: igual, thermia: temperatura; que é como chamamos os animais que possuem temperaturas corporais constantes). Haemothermia, por sua vez era relacionado evolutivamente com o Crocodylia (crocodilos, jacarés e gaviais) formando o grupo Thecodontia. Tartarugas, cágados e jabutis (Chelonia) eram mais aparentados com Thecodontia (Crocodylia+(Aves+Mammalia)). Por fim, o grupo mais basal de Amniota era Lepidosauria, o qual abrange tuataras, serpentes e lagartos (Figura 1).
Fig. 1. Relações entre as cinco assembleias de amniotas viventes defendida por Gardiner (1982; ver também Lovtrup, 1985) modificado de Gauthier e colaboradores (1988)
O trabalho experimental de Gauthier
Os resultados de Gardiner se baseam exclusivamente em animais viventes, conforme já foi dito e ecoaram por quase uma década. Lovtrup (1985) publicou um trabalho no qual afirmou ter encontrado maior suporte à hipótese de Gardiner. Até que, em 1988, um célebre manuscrito chegou para revolucionar a visão que a comunidade científica da época tinha sobre os dados paleontológicos. Gauthier e seus colegas (1988), baseados na hipótese de filogenia dos amniotas proposto por Gardiner, publicaram o primeiro estudo demonstrando empiricamente (ou seja, através de experimentos práticos e não calcados apenas em conjecturas) que os fósseis poderiam contribuir e muito na elucidação das hipóteses de parentesco dos seres vivos atuais. Para isso, eles conduziram uma nova análise, só que dessa vez incluindo espécies extintas, obtendo uma árvore marcadamente diferente e depois a submeteram a alguns testes para confirmar sua validade. Para entendermos como os autores chegaram a suas conclusões precisamos compreender os experimentos que a equipe realizou. Gauthier partiu de uma pergunta: os Fósseis poderiam alterar as nossas hipóteses de parentesco entre as biotas recentes? Para responder isso, ele conduziu um estudo em três etapas.
(1) Primeiramente, antes de rodar uma nova análise incluindo os fósseis, os pesquisadores destrincharam e reviram todas as características utilizadas por Gardiner e perceberam que havia alguns equívocos e erros na interpretação dos caracteres. Gardiner afirmara, por exemplo, que um coração dividido em quatro câmaras é homólogo (mesma origem; para melhor entendimento desta terminologia veja aqui) em crocodilos, aves e mamíferos, o que reforçava a relação de parentesco entre eles. Contudo, quando acompanhamos o desenvolvimento embrionário do septo interventricular destes animais, vemos que nos mamíferos ele se desenvolve a partir de uma crista de tecido endocárdico na parede de trás do ventrículo, enquanto que nas aves e nos crocodilos o septo surge de varias protuberâncias musculares pouco recobertas por endocárdio na lateral do ventrículo. Apesar de serem estruturas semelhantes nos adultos, elas não possuem a mesma origem no embrião, ou seja, não são homólogas. Logo, os equívocos nas interpretações de Gardiner o levaram a estabelecer homologias entre mamíferos e aves que não condiziam com a realidade.
(2) Depois de corrigir a lista de características, Gauthier rodou duas análises, uma contendo apenas os dados morfológicos dos cinco grupos viventes de Gardiner e outra incorporando 29 espécies de amniotas extintos. Foi então que uma nova hipótese começou a tomar forma.
A análise com dados dos animais viventes
A árvore filogenética obtida utilizando apenas os cinco grupos de amniotas viventes se assemelhou à hipótese clássica de Gardiner de 1982, com apenas uma pequena diferença: os crocodilos e não os mamíferos eram mais relacionados com as aves, trazendo dúvidas quanto a validade do antigo grupo Haemothermia (Ver figura 2). Isso implica em inferências importantíssimas para nossa compreensão da evolução da homeotermia. Primeiro, que um jacaré é o parente mais próximo das aves do que qualquer outro animal vivo hoje. Além disso, muitas características que antes eram vistas como homólogas entre aves e mamíferos e que estariam presentes no suposto ancestral comum destas duas linhagens, na verdade surgiram duas vezes independentemente na árvore da vida dos vertebrados. Talvez em resposta provavelmente as mesmas pressões evolutivas.
Fig. 2. Árvore obtida da análise de Gauthier (1988) apenas com amniotas viventes. Note que ela se assemelha à árvore anterior de Gardiner (1982), contudo as aves estão mais relacionadas com crocodilos do que com os mamíferos.
A análise com dados combinados (animais viventes + extintos)
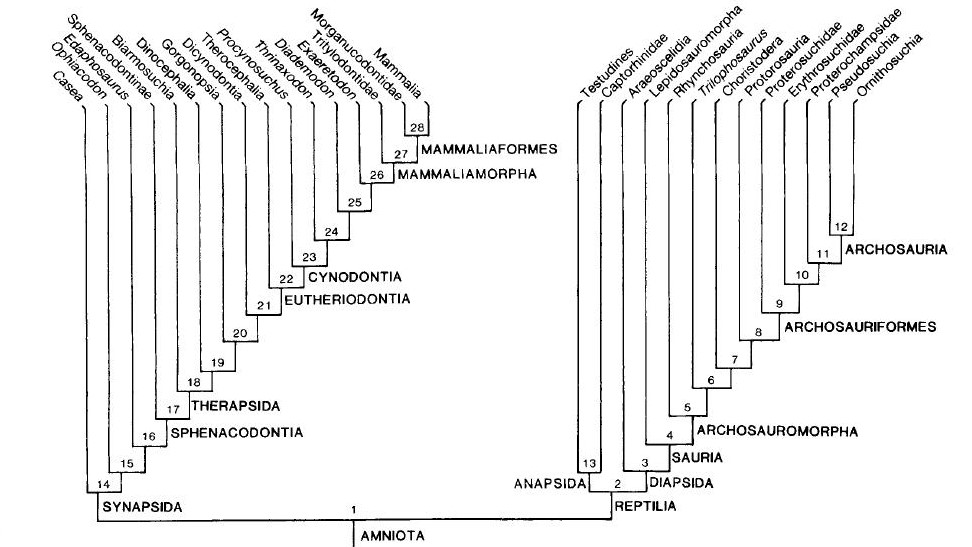
Na segunda análise na qual Gauthier acrescentou os fósseis, uma árvore completamente diferente surgiu (ver figura 3). Não só aves e crocodilos estavam agrupados juntos, mas lagartos e serpentes (Lepidosauria) agora estavam mais relacionados a Aves + Crocodylia do que as tartarugas. O mais estranho foi que os mamíferos agora estavam na base da árvore, formando uma grande dicotomia inicial: Mamíferos e todos seus parentes extintos de um lado versus lepidossauros, quêlonios, crocodilos, aves e todos seus parentes extintos relacionados do outro. Ao primeiro grupo, chamamos de Synapsida (amniotas que apresentam uma fenestra temporal no crânio) e ao segundo, Reptillia (que compreende tanto amniotas que possuem duas ou nenhuma fenestra temporal, Diapisida e Anapsida, respectivamente). Contudo, não bastava obter uma árvore completamente diferente, ela precisava ser mais bem justificada.
Fig. 3. Árvore obtidida por Gauthier (1988) combinando tanto espécies fósseis quanto viventes. Note que há uma dicotomia basal, separando mamíferos e seus parentes extintos (Synapsida) de um lado versus todos os répteis atuais + Aves (Reptilia) do outro.
Análises posteriores – Esmiuçando a nova hipótese obtida
Na última etapa do estudo (3), uma série de experimentos computacionais foram conduzidos por Gauthier, alguns serão explicados mais adiante e que culminaram em um achado esperançoso, principalmente para nós paleontólogos: Fósseis são fundamentais para elaboração das hipóteses de relação entre as espécies viventes. Gauthier e seus colegas queriam saber o porquê que as árvores diferiram tanto nas análises com e sem os fósseis. Para isso, eles fizeram algumas análises posteriores, contudo só irei detalhar três delas neste texto que acredito serem mais interessantes para nossa discussão. Primeiro, os autores compararam o índice de consistência de sua nova árvore com o índice da árvore de Gardiner. Este índice mostra o quão robusto e conciso estão seus resultados, no caso, sua hipótese de evolução de uma linhagem. O interessante foi que mesmo adicionando várias espécies fósseis na sua análise, Gauthier encontrou um valor de índice de consistência semelhante ao de Gardiner, mostrando que mesmo os fósseis alterando drasticamente a hipótese, a árvore continuava tão confiável quanto uma utilizando apenas espécies viventes.
Numa análise posterior, Gauthier e colaboradores removeram todos os grupos fósseis do lado “sinapsídeo” da árvore, deixando apenas os animais viventes desse ramo (mamíferos) junto com todos do lado “Reptillia” (viventes e extintos) e rodaram a análise. Paralelamente, foi feito o oposto, todas as linhagens extintas de Reptillia foram retiradas deixando apenas seus representantes viventes e o lado “sinapsídeo” da árvore (viventes e extintos) e rodaram a análise. Os autores viram que não importava qual fóssil de Reptillia fosse retirado, a árvore final não se alterava. Porém, quando os fósseis de sinapsídeos eram retirados, a árvore adquiria o novo padrão proposto por Gardiner (1982; Figura 3).
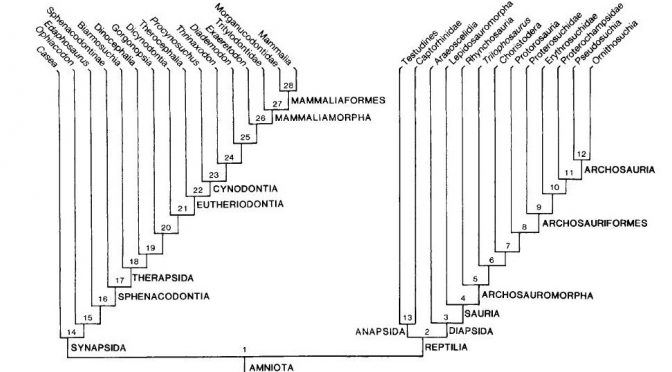
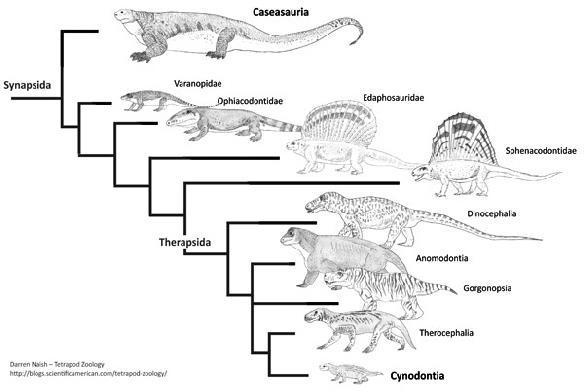
Gauthier então percebeu que os principais responsáveis pela nova topologia da árvore eram os fósseis de sinapsídeos. Mas por que isso? De acordo com os autores, quanto mais antiga é a origem de uma linhagem e quanto mais derivada for a morfologia de seus representantes atuais (como é o caso dos mamíferos), mais os fósseis serão importantes para elucidar sua evolução. Basta compararmos os mamíferos atuais com seus parentes extintos, os “pelicossauros” como Casea, Ophiacodon, Edaphosaurus, Sphenacodon (figura 4) para ver o “abismo morfológico” que separam essas linhagens. Quando observamos os mamíferos atuais, dificilmente conseguimos relaciona-los com outro grupo de animal vivente e quando tentamos, certamente estaremos fadados ao erro, assim como Gardiner, em 1982 equivocadamente agrupou Aves e Mammalia no antigo grupo Haeomothermia. Porém, quando olhamos para os fósseis vislumbramos um mundo completamente “novo”. As formas extintas possuem combinações únicas de características basais e derivadas que frequentemente se apresentam numa serie gradual de mudanças, que nos permitem acompanhar suas histórias evolutivas de maneira tal que possamos observar espécies diferentes ficando cada vez mais semelhantes conforme voltamos no tempo. Os fósseis são uma das poucas evidências diretas da evolução e é por isso, que os dados paleontológicos se tornam tão fundamentais para elucidar as relações evolutivas dos grupos viventes. Principalmente, quando o objeto de estudo são animais tão diferentes como seu cachorro e um pardal na janela de casa.
Outro experimento que a equipe de Gauthier conduziu consistiu basicamente em retirar todas as linhagens viventes da matriz de características e fazer uma nova análise. Apesar de ser um experimento simples, o resultado obtido foi revelador: a nova hipótese representada na figura 3 veio à tona novamente, porém sem os animais viventes. Este resultado, associado à análise inicial das características utilizadas no trabalho de Gardiner (1982), derruba os argumentos dos críticos a respeito da incompletude do registro fóssil. De fato a informação que provem dos fosseis é mais incompleta do que as que podemos retirar dos animais viventes, mas mesmo assim os fósseis estão longe de serem menos informativos em uma análise filogenética. Gauthier demonstrou isso quando retirou todas as espécies viventes da análise e mesmo assim alcançou a nova hipótese. Além disso, Gauthier enfatizou que incompletude não é exclusiva de fósseis. Ela pode ocorrer naturalmente nos animais. Quatro das características utilizadas na análise de Gardiner eram do osso quadrado, um osso na base do crânio dos vertebrados no qual se articula a mandíbula. Contudo, nos sinapsídeos mais derivados, os cinodontes (o qual mamíferos fazem parte), esse osso foi gradativamente sendo reduzido e alocado cada vez mais para trás do crânio junto com uma série de ossos da mandíbula. Esta condição alcançou o extremo nos mamíferos, onde o osso quadrado e um grupo de ossos que uma vez pertenceram à mandíbula, se reduziram e modificaram sua função ao ponto de formarem o que hoje são nossos ossículos do ouvido médio (o quadrado dos outros vertebrados é a bigorna nos mamíferos). Logo, aquelas quatro características baseadas na morfologia do osso quadrado, não se aplicam aos mamíferos, pois o quadrado deles se alterou tanto que fica difícil comparar com os outros animais, ou seja, este é um dado naturalmente faltante. Além disso, Gauthier percebeu que Casea (o fóssil mais basal e antigo de sinapsídeo e que consequentemente esperávamos maior incompletude) apresentava 26% de informação faltando, enquanto que os mamíferos atuais tinham em média 15% de dados morfológicos faltando ou difíceis de interpretar. Esses 11% de diferenças são realmente significantes? Gauthier não só provou que não há diferença significante como também nos mostrou que informação incompleta não é exclusiva dos fósseis.
Fig. 4. Outra árvore filogenética, dessa vez apenas com sinapsídeos. Note a variedade de formas desde as mais basais, como Casesauria (um “Pelicossauro”), até os parentes mais próximos dos mamíferos, um cinodonte não-mamífero.
A Renascença dos fósseis
Após demonstrar empiricamente quão importante os fósseis são para nossa compreensão da evolução das linhagens viventes, Gauthier deu um solavanco nos sistematas, convidando-os a se debruçarem no assunto. Desde então, estudos importantes vêm sendo conduzidos, cada vez mais enaltecendo o uso de dados paleontológicos em reconstruções filogenéticas e desenvolvendo metodologias para minimizar o efeito da incompletude dos dados de organismos tanto viventes quanto extintos, o famigerado missing data que os cladistas tanto abominam (veja Donoghue et al., 1989; Smith, 1998; Wilkinson; Benton, 1995). Exemplos como estes de Gardiner e Gauthier nos mostram como a Ciência é dinâmica. O que antes era tido como verdade e que hoje é obsoleto, não necessariamente deixa de ser útil. Se Gardiner não tivesse se aventurado nas relações de Amniota, Gauthier e toda uma geração de sistematas não seriam impulsionadas a refletir sobre o assunto e talvez o reconhecimento da importância dos fósseis na reconstrução da evolução da vida na Terra poderia ser ainda mais postergada. Por fim, quando ignoramos as informações contidas no registro fóssil, estamos ferindo o princípio da Evidência Total (um tema que será abordado em postagens futuras do blog). Este princípio pode ser exemplificado com uma ótima analogia que um estimado amigo uma vez me fez: não usar os dados paleontológicos para inferir filogenia é como ter um bebê de colo e joga-lo pela janela só porque ele não é um adulto.
Referências:
AX, Peter. 1987. The phylogenetic system: the systematization of organisms on the basis of their phylogenesis. DARWIN, C. A Origem das Espécies. Hemus – Livraria Editora Ltda, São Paulo, SP.
DONOGHUE, Michael J. 1989. Phylogenies and the analysis of evolutionary sequences, with examples from seed plants. Evolution, v. 43, n. 6, p. 1137-1156.
GARDINER, BRIAN G. 1982. Tetrapod classification. Zoological Journal of the Linnean Society, v. 74, n. 3, p. 207-232.
GAUTHIER, Jacques; KLUGE, Arnold G.; ROWE, Timothy. 1988. Amniote phylogeny and the importance of fossils. Cladistics, v. 4, n. 2, p. 105-209. HENNIG, Willi. 1950. Grundzuge einer Theorie der phylogenetischen Systematik.
LOVTRUP, Soren. 1985. On the classification of the taxon Tetrapoda. Systematic Zoology, v. 34, n. 4, p. 463-470.
PATTERSON, Colin. 1981. Significance of fossils in determining evolutionary relationships. Annual Review of Ecology and Systematics, v. 12, n. 1, p. 195-223.
SMITH, Andrew B. 1998. What does palaeontology contribute to systematics in a molecular world?. Molecular phylogenetics and evolution, v. 9, n. 3, p. 437-447.
WILKINSON, Mark; BENTON, Michael J. 1995. Missing data and rhynchosaur phylogeny. Historical Biology, v. 10, n. 2, p. 137-150.
Geovane Alves de Souza, Graduado em licenciatura e bacharelado em Ciências Biológicas pela Universidade Estadual de Londrina. Atualmente é mestrando em Zoologia pelo Museu Nacional/UFRJ. Já desenvolveu pesquisas na área de parasitologia de animais silvestres, hoje conduz estudos de osteohistologia com titanossauros.
Há algumas semanas, um estudo publicado na prestigiosa revista ‘Nature’ chamou a atenção do mundo e veio abalar as estruturas de um consenso secular na paleontologia de dinossauros. O estudo publicado na revista Nature foi desenvolvido por Baron (Universidade de Cambridge) e colaboradores, e contou com uma amostragem abrangente de espécies basais de dinossauros e outros Dinosauromorpha.
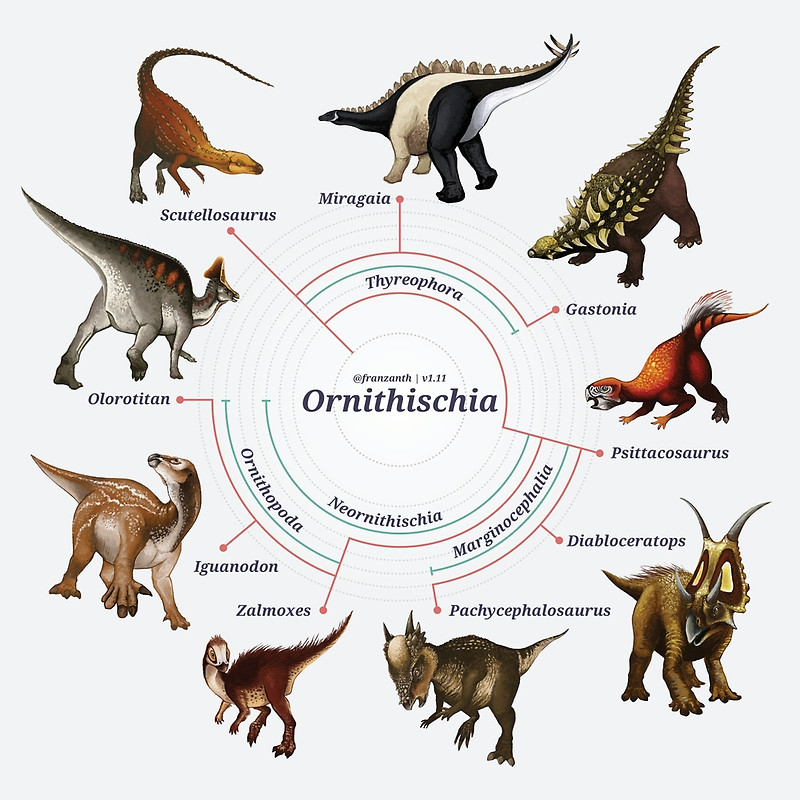
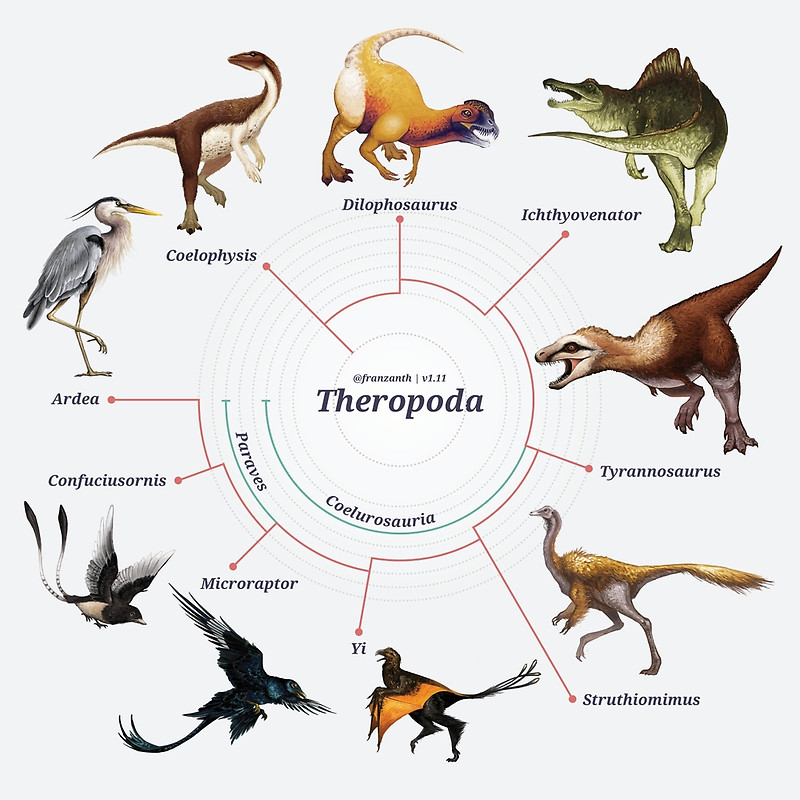
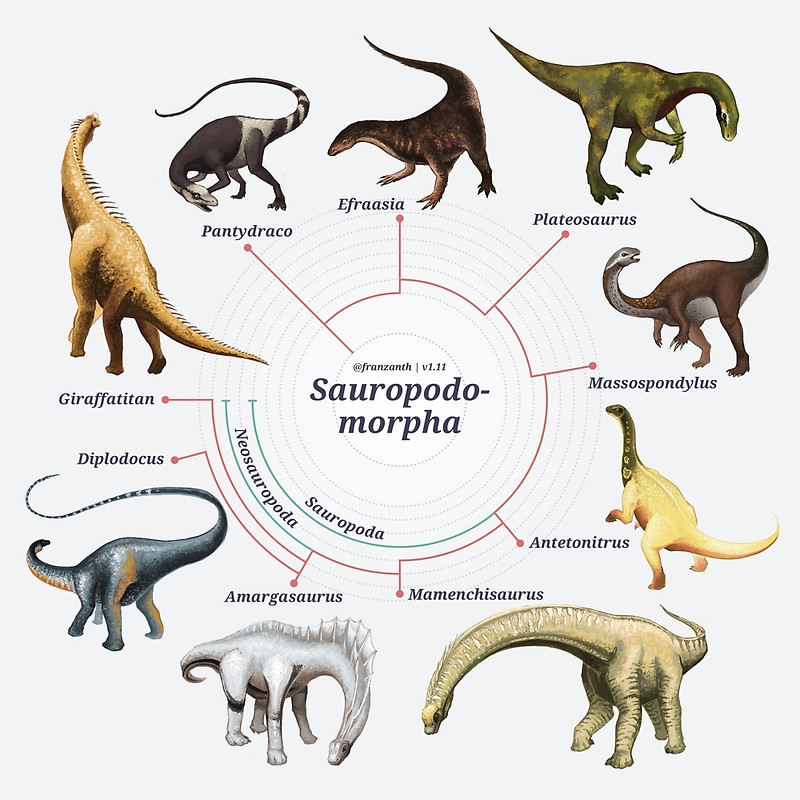
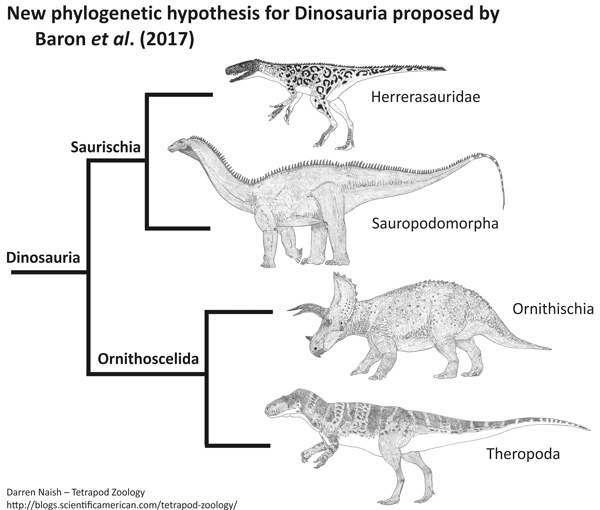
A novidade do estudo de Baron e colegas é que uma nova topologia para a árvore evolutiva dos dinossauros foi obtida, onde os dinossauros ornitísquios (que incluem desde os estegossauros até os dinossauros “bico de pato”, veja imagem a seguir) caem como grupo irmão dos dinossauros terópodes (grupo de dinossauros que inclui basicamente todos os dinossauros carnívoros), formando um clado denominado de ‘Ornithoscelida’. Esse resultado altera completamente o consenso tradicional sobre a evolução dos dinossauros, que colocava os dinossauros terópodes e sauropodomorfos (grupo dos dinossauros herbívoros de pescoço e cauda longa) juntos, formando o clado clássico conhecido como Saurischia.
Dinossauros ornitísquios. Arte de Franz Anthony (http://franzanth.com/).
Dinossauros terópodes. Arte de Franz Anthony.
Dinossauros sauropodomorfos. Arte de Franz Anthony.
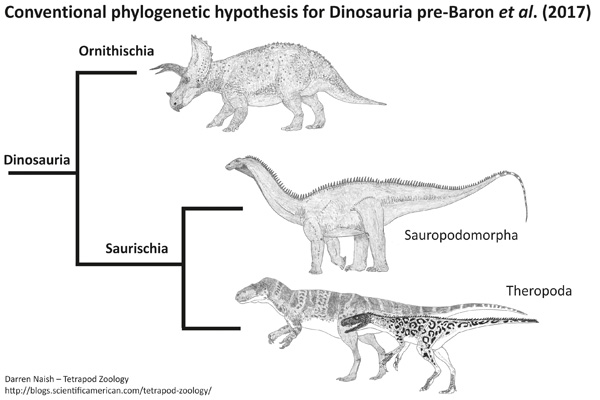
Cladograma de acordo com a filogenia clássica ou mais convencional dos dinossauros. Imagem por Darren Naish.
Cladograma ilustrando a filogenia proposta por Baron et al. (2017), separando os dinossauros terópodes e sauropodomorfos e sustentando o clado denominado de ‘Ornithoscelida’ (Theropoda + Ornitischia). Imagem por Darren Naish.
A filogenia tradicional dos dinossauros, que se sustenta há quase 130 anos, sempre partiu do princípio de que dinossauros terópodes e sauropodomorfos formavam um grupo monofilético, ou seja, que consistiam de um agrupamento verdadeiro, que reunia uma espécie ancestral e todos os seus descendentes.
Apesar do trabalho de Baron desafiar a proposta convencional das relações evolutivas dos dinossauros, o ordenamento que ele propõe em seu artigo não é muita novidade. Propostas alternativas, incluindo essa de Ornithoscelida, sempre existiram e foram consecutivamente testadas ao longo do tempo. O que acontece é que, nos últimos anos, novas espécies de dinossauros basais foram descobertas e descritas e pudemos ter acesso a novas informações sobre como se deu o seu processo evolutivo de certos aspectos morfológicos dos dinossauros. Isso deu mais resolução à nossa compreensão sobre a evolução desse grupo. O que Baron fez foi reunir essa informação em uma ampla matriz de dados morfológicos e testá-la. O resultado foi que, com a nova amostragem de táxons basais de dinossauros, o arranjo filogenético que melhor explica o que observamos é a união de Theropoda e Ornithischia em um mesmo grupo: Ornithoscelida.
As grandes propostas alternativas sobre a evolução de Dinosauria.
Já posso queimar toda a minha bibliografia sobre dinossauros? Foto de Darren Naish.
Não preciso nem dizer que isso causou um reboliço na paleontologia e uma acalorada discussão entre paleontólogos, né?
Mas, calma, você não precisa sair por aí queimando todos os seus livros sobre dinossauros. Toda nova proposta que muda drasticamente uma ideia merece ser testada e reavaliada antes de definitivamente adotada.
Com a ascensão desse trabalho, vários paleontólogos do mundo todo se reuniram para analisar minuciosamente a matriz de dados morfológicos usada por Baron e colegas. Há muito tempo precisávamos de uma amostragem abrangente, incluindo mais táxons basais de Ornithischia e isso o trabalho de Baron tem de positivo! Porém, onde aparentemente o trabalho de Baron falha, é na matriz de dados em si. Muitos colegas paleontólogos do mundo têm apontado falhas na codificação da matriz filogenética apresentada no estudo publicado na Nature, e eles já estão trabalhando em uma réplica. Nos próximos meses, uma publicação reunindo paleontólogos de diversas nacionalidades deverá ser publicada reavaliando a matriz de dados de Baron.
“Alegações extraordinárias exigem evidências extraordinárias” Carl Sagan
A hipótese de Ornithoscelida não pode ser totalmente desconsiderada, já que a topologia filogenética obtida por Baron e colaboradores explica muito bem a distribuição de alguns caracteres morfológicos em dinossauros. O caracter mais claro para exemplificar essa questão talvez seja a presença de estruturas tegumentárias (i.e. penas e estruturas similares à penas) tanto em Ornithischia quanto em Theropoda. Estruturas as quais ainda não foram encontrada em fósseis de Sauropodomorpha (o que não significa que eles definitivamente não as possuíam!!!).
Fóssil de Psittacosaurus, um dinossauro Ornithischia, com estruturas tegumentárias possivelmente homólogas às penas encontradas em dinossauros terópodes.
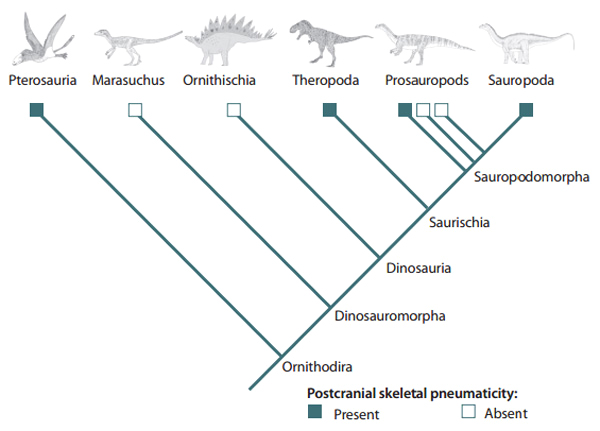
Alguns paleontólogos argumentam que, apesar de explicar bem a distribuição de alguns caracteres, essa proposta filogenética não sustenta tão bem outras questões anatômicas muito importantes, como a pneumaticidade nos ossos, observada tanto em Theropoda como em Sauropodomorpha – mas não em Ornithischia -, ou mesmo o clássico formato do quadril trirradiado.
A questão, pelo visto, continuará sendo quais caracteres morfológicos evoluíram independentemente ou não…
Presença/ausência de pneumaticidade óssea em Ornithodira.
Pneumticidade em vértebra de um dinossauro terópode. Característica também presente em Sauropodomorpha.
Outra grande crítica ao trabalho de Baron foi que a sua proposta filogenética aponta a origem dos dinossauros como sendo europeia, o que para a grande maioria dos paleontólogos não faz sentido algum.
A lição maior do trabalho de Baron é que ainda temos muito o que investir no estudo de dinossauros basais. Vamos aguardar o trabalho sobre a revisão da matriz de dados que sustenta Ornithoscelida ser publicado e torcer para novas descobertas de dinossauros basais sejam feitas!
Assista o vídeo sobre essa questão em nosso canal. Entrevistamos o paleontólogo argentino Diego Pol, um dos especialistas envolvidos na re-avaliação da matriz de dados do trabalho de Baron:
Baron, M. G., Norman, D. B. & Barrett, P. M. 2017. A new hypothesis of dinosaur relationships and early dinosaur evolution. Naturedoi:10.1038/nature21700