Texto por Mário G. F. Esperança Júnior
As paisagens que reconhecemos à nossa volta possuem uma história bastante dinâmica. Por exemplo: rios mudam de curso, lagos secam, mares retraem e avançam sobre os continentes… Em um intervalo de tempo mais amplo, montanhas são formadas e erodidas, oceanos se fecham, novas espécies de organismos surgem enquanto outras são extintas. Evidências geológicas mostram que esses processos são recorrentes e se sucederam por todo o planeta desde seus primórdios.
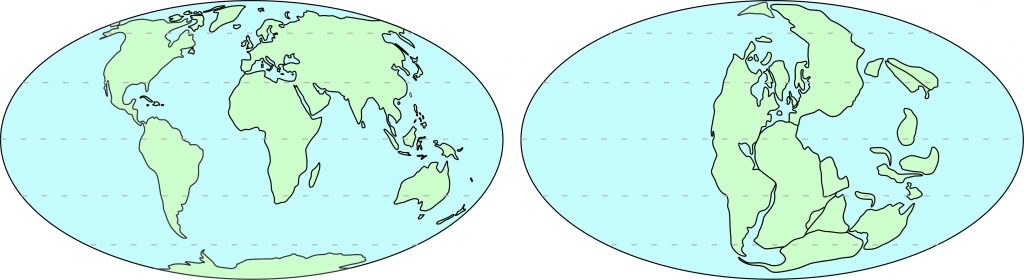
No sul do Brasil, afloram rochas do período Permiano, intervalo que compreende de 298 a 252 milhões de anos atrás. Essas rochas são ricas em fósseis da extinta Flora Glossopteris, plantas as quais também são encontradas em depósitos sedimentares da África, Antártida, Austrália e Índia. Atualmente, estas regiões estão separadas por extensos oceanos, que são barreiras intransponíveis para grande parte dos organismos terrestres, incluindo as plantas. Sendo assim, pode-se deduzir que tais áreas estiveram unidas durante o Permiano, e mais tarde se afastaram até chegarem em suas localizações atuais. Dessa forma, as floras e faunas que antigamente colonizaram regiões contíguas, passaram a ser encontradas na forma de fósseis em lugares distantes.
A evolução desse pensamento científico levou muitos anos até chegar aos moldes que hoje conhecemos. Mas para entendermos a sucessão dos fatos que nos levam a resposta acerca de tais semelhanças fossilíferas, precisamos voltar alguns séculos atrás…
As primeiras ideias
Com a chegada dos europeus ao continente americano no século XV e com a confecção dos primeiros mapas em escala global, notou-se que a costa oeste da África e leste da América do Sul possuem contornos semelhantes, assim sugerindo que tais continentes estivessem em algum momento unidos. Àquela época, contudo, perduravam as ideias bíblicas de que a Terra seria jovem, e o conhecimento científico limitava-se à observação direta do meio devido à ausência de tecnologia capaz de comprovar tais fenômenos. Somente no século XVII é que surgiram as primeiras evidências acerca de uma idade para a Terra. Ao observar intrusões de granitos, bem como o ciclo de erosão e deposição de sedimentos, o naturalista escocês James Hutton deduziu que havia muito tempo envolvido nesses processos, concluindo que a Terra seria muito antiga, “sem vestígio de um começo, nem perspectiva de um fim” (Hutton, 1788). Décadas mais tarde, o matemático irlandês Lord Kelvin calculou a idade do planeta, que estaria entre 20 e 400 milhões de anos, dado o presente equilíbrio térmico e fluxo de calor da superfície. Esse cálculo foi posteriormente refinado, mas somente no final do século XIX, com a descoberta da radioatividade por Henri Becquerel e o subsequente desenvolvimento das técnicas de datação radiométricas, é que se chegou à atual idade de 4,56 bilhões de anos. Essa escala de tempo é grande o suficiente para explicar processos naturais muito lentos para a humanidade, porém recorrentes na história da Terra.
A comprovação da existência de um tempo profundo talvez tenha sido um dos principais fatores para o estabelecimento de uma teoria que explicasse o encaixe dos continentes percebido séculos antes. Tais ideias foram fomentadas no início do século XX com o trabalho do geólogo austríaco Eduard Suess (Fig. 1), ao notar a presença das folhas fósseis do tipo Glossopteris na América do Sul, África e Índia. A coocorrência das mesmas plantas em áreas distantes e separadas por oceanos é um indicativo de que essas massas de terra já estiveram conectadas, por fim denominando essa antiga região como Terra de Gondwana (Suess, 1885). A fim de explicar tal observação, Suess deduziu que o resfriamento do planeta causaria o adensamento de certas áreas que consequentemente afundariam formando os oceanos. Entre essas regiões, haveria pontes de terra permitindo o intercâmbio da fauna e flora, sendo que estas conexões mais tarde submergiriam resultando no padrão geográfico atual. Essa teoria, no entanto, não foi bem aceita por não existirem evidências físicas de tais fenômenos. Nos anos subsequentes, propuseram-se, ainda, que uma massa de terra primária poderia ter sido partida em continentes menores devido à expansão volumétrica do planeta (Mantovani, 1889), e também que a movimentação das mesmas seria fruto da força das marés – hipóteses que foram rejeitadas por físicos e matemáticos.

Foi assim que, em 1915, o meteorologista alemão Alfred Wegener propôs que as massas continentais outrora estivessem amalgamadas no supercontinente Pangeia (Fig. 2), dadas semelhanças não somente entre os fósseis, mas também com as rochas e estruturas geológicas encontradas nos dois lados do Atlântico (Wood, 1980). Por exemplo, a presença de fósseis de Mesossauros, antigos répteis marinhos encontrados na América do Sul e África, também apontavam para tal proximidade. Além disso, a ampla presença de rochas glaciais, denominadas de tilitos, sugeria uma união de diversas áreas próximo ao polo sul. Com essas e outras evidências, propôs-se a teoria do afastamento, ou deriva dos continentes, fenômeno que seria decorrente da rotação da Terra (Wegener, 1915). Contudo, a explicação de Wegener sobre o motor responsável pela deriva não foi bem aceita pela comunidade científica. O verdadeiro mecanismo responsável pela afastamento dos continentes só foi estabelecido anos mais tarde pelo geólogo inglês Arthur Holmes. Conforme a ideia apresentada, existiriam fluxos convectivos verticais abaixo dos continentes, constantemente alimentados pelo calor gerado por elementos radioativos do interior da Terra. Tais fluxos arrastariam as massas continentais continuadamente, ocasionando na formação de assoalho oceânico nas regiões em que as correntes quentes sobem, e na criação de montanhas na borda em que o fluxo frio desce (Holmes, 1931). Entretanto, as evidências do processo descrito por Holmes surgiram somente anos depois com a ampliação do conhecimento sobre o fundo oceânico.
Nesse sentido, o conhecimento a respeito da Dorsal Mesoatlântica exerceu um papel fundamental no entendimento da deriva continental. Esta cadeia de montanhas submarina foi inicialmente identificada em 1872, em uma expedição liderada pelo naturalista escocês Charles W. Thomson com o objetivo de investigar a área por onde passariam cabos submarinos de telégrafo, ligando a Europa à América do Norte. Mas foi somente após a Segunda Guerra Mundial que realmente se intensificaram as pesquisas sobre o assoalho marinho, o que incluiu os primeiros mapeamentos do fundo oceânico realizados por sonar (Heezen et al., 1959). Com isso, constatou-se que a Dorsal Mesoatlântica, em quase sua totalidade, inclui um vale em rifte – regiões de vulcanismo intenso em que terremotos são frequentes. Além disso, datações do assoalho marinho mostraram que o mesmo é composto por rochas vulcânicas jovens. Dessa maneira, a formação da litosfera oceânica é o resultado da atividade vulcânica recente desses riftes, que pouco a pouco afastam porções mais antigas, expandindo o oceano (Heezen, 1960; Dietz, 1961). Assim, ao menos parte das ideias de Holmes já possuía alguma comprovação no início da década de 60.
O nascimento de uma teoria unificadora
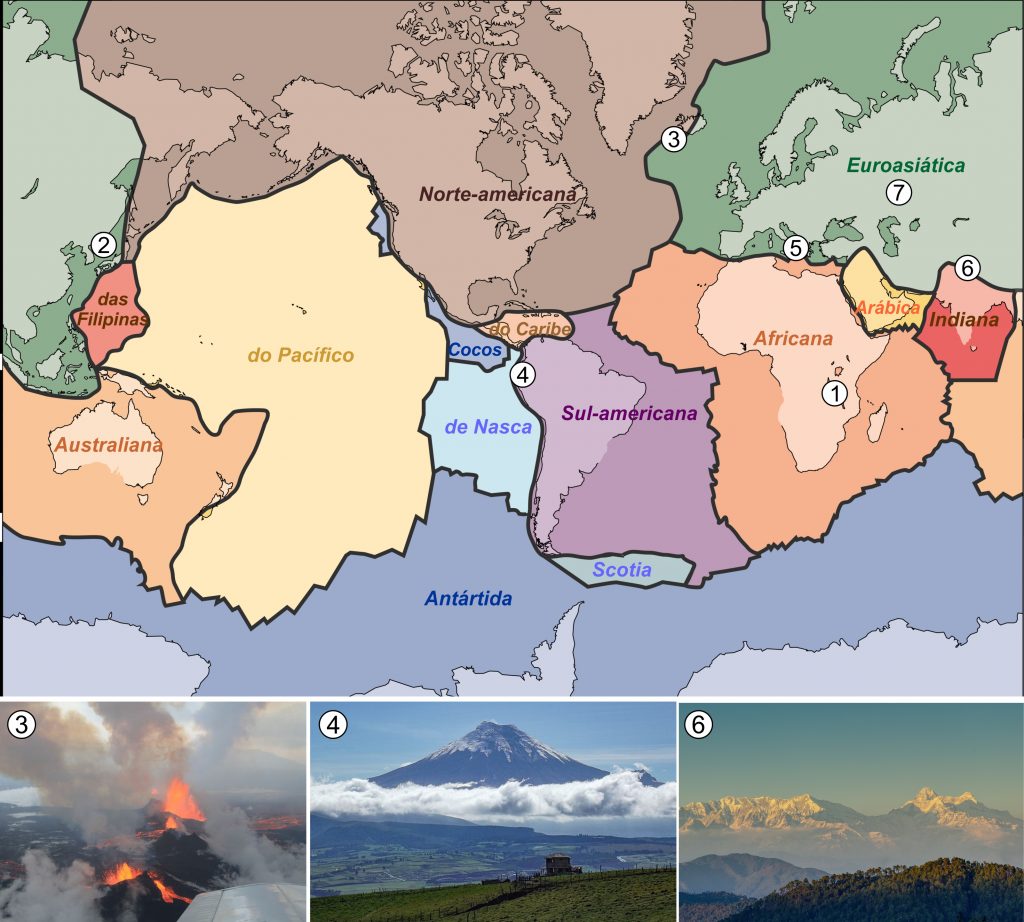
No entanto, existia uma interessante questão em aberto: já que o assoalho dos oceanos é criado ao longo das dorsais, haveria algum mecanismo capaz de destruí-lo? Com o avanço do mapeamento do fundo marinho durante a década de 60, o geólogo canadense John Tuzo Wilson constatou que certas falhas geológicas, dorsais e fossas submarinas se interconectam formando grandes placas. Assim, a litosfera seria dividida em diversas placas rígidas que se movimentam e submergem sob os arcos vulcânicos ao envelhecerem e resfriarem. À medida que o oceano é destruído, os continentes circundantes se aproximam e eventualmente colidem formando cadeias de montanhas, a exemplo dos Himalaias (Wilson, 1963; 1965; 1966). Dessa forma, a abertura e fechamento dos oceanos seria um encadeamento cíclico, e portanto, repetitivo ao longo da história da Terra (Fig. 3). Mais tarde, esse processo foi denominado, em sua homenagem, de Ciclo de Wilson – uma importante contribuição ao que hoje conhecemos por Teoria da Tectônica de Placas (Fig. 4).
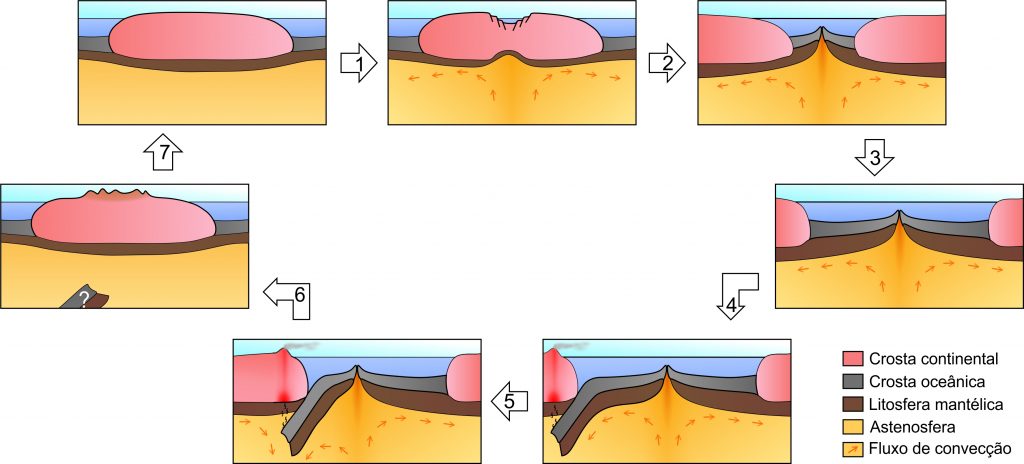
Atualmente, o avanço tecnológico permite inferir temperaturas para o interior da Terra através de tomografia sísmica. Com isso, comprovou-se a existência de regiões quentes próximas às dorsais, enquanto que as temperaturas são mais baixas onde antigas placas oceânicas afundaram. Essa ascensão de material quente e descida de rochas frias demonstra o processo de convecção proposto por Holmes no começo do século passado. O desenvolvimento da tecnologia permitiu, também, mensurar com grande precisão a taxa de afastamento dos continentes (e portanto, a taxa de expansão do assoalho oceânico). No caso do Atlântico, o afastamento varia de 1,8 centímetro por ano, próximo à Islândia; até 3,5 centímetros ao ano, entre o Brasil e o sul da África (Grotzinger e Jordan, 2013), velocidades realmente lentas para a percepção da humanidade. Entretanto, distâncias consideráveis podem ser alcançadas, levando-se em consideração o amplo intervalo de tempo que compreende a fascinante história da Terra.
Referências
Dietz, R.S. 1961. Continent and ocean-basin evolution by spreading of the sea floor. Nature 190, 854-857.
Grotzinger J., Jordan T.H. 2013. Para entender a Terra. 6. ed. Porto Alegre: Bookman.
Heezen, B.C., Tharp, M., Ewing, M. 1959. The floors of the ocean, I. The North Atlantic. Geological Society of America, Special Paper 65.
Heezen, B.C. 1960. The rift in the ocean floor. Scientific American 203, 98-110.
Holmes, A. 1931. XVIII. Radioactivity and Earth Movements. Transactions of the Geological Society of Glasgow 18(3), 559–606.
Hutton, J. 1788. X. Theory of the Earth; or an Investigation of the Laws observable in the Composition, Dissolution, and Restoration of Land upon the Globe. Earth and Environmental Science Transactions of The Royal Society of Edinburgh 1(2), 209 – 304.
Mantovani, R. 1889. Les fractures de l’écorce terrestre et la théorie de Laplace. Bulletin de la Société des sciences et arts de l’Ile de la Réunion, 41–53.
Suess, E. Das Antlitz der Erde. 1885. Leipzig: Freytag.
Wegener, A. 1915. Die Entstehung der Kontinente und Ozeane. Braunschweig: Vieweg.
Wilson, J.T. 1963. A possible origin of the Hawaiian islands. Canadian Journal of Physics 41, 863-870.
Wilson, J.T. 1965. A new class of faults and their bearing on continental drift. Nature 207, 343-347.
Wilson, J.T. 1966. Did the Atlantic close and then re-open? Nature 211, 676-681.
Wood, R.M. 1980. Coming Apart at the Seams. New Scientist 85, 252-254.