>
Durante a Era Mesozóica, os mares eram habitados por uma formidável diversidade de répteis marinhos. O enfoque desse texto introdutório é para os três grupos extintos mais conhecidos desses animais: os Ictiossauros, os Plesiossauros (Pliossauros inclusos) e os Mosassauros.

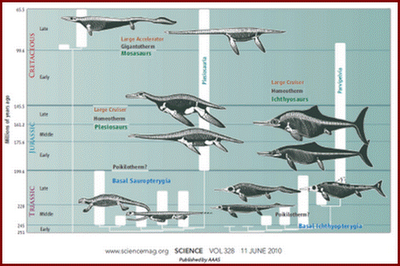
Figura 1: Cladogramas de répteis marinhos mesozóicos.
Fonte: SCIENCE. www.siencemag.org

Figura 2 Reconstrução de Ictiossauro caçando um amonite.
By Jorge Gonzales.
O primeiro grupo a ser apresentado, os Ictiossauros (figura 2), eram extremamente adaptados à vida marinha. Possuiam um corpo aquadinâmico, semelhante ao de um golfinho, com membros em formatos de pás, nadadeiras caudais verticais, como as de um tubarão, focinhos longos, e, em espécies mais derivadas, barbatanas dorsais (há restos de fósseis com registro de contorno de barbatanas dorsais para alguns gêneros, como observou McGowan à respeito dos melhores fósseis encontrados para esse grupo na região de Holzmaden, na Alemanha).
A locomoção dos Ictiossauros se dava por propulsão em alta velocidade dada por meio de sua poderosa cauda com o direcionamento da barbatana (para os indivíduos que tinham essas características já desenvolvidas) e nadadeiras. O seu hábito alimentar envolvia basicamente uma dieta de cefalópodes, ingeridos por meio de sucção (como algumas baleias modernas). Sua reprodução era ovovivípara, ou seja, os filhotes eram expelidos do corpo já formados: há amplo registro fóssil de fetos associados as mães que teriam morrido por complicações no parto — Para alguns pesquisadores, todavía, há também a teoria que os fetos poderiam ser expelidos do corpo das mães pós-morten, como McGowan propôs baseado em carcaças de baleias na Tasmania que liberaram os fetos depois de mortos.
Os Ictiossauros surgiram no início do Período Triássico e extiguiram-se no Neocretáceo, há cerca de 90 milhões de anos. Um dos gêneros encontrados na Colômbia, Platypterygius, foi um dos últimos antes que o grupo se extiguisse. O gênero Platypterygius encontrava-se também nos mares da América do Norte, Europa, Rússia, Índia e Austrália.

Figura 3: Reconstrução computadorizada de um Plesiossauro.

Figura 4: Reconstrução computadorizada de um Plesiossauro Elasmossauro. Fonte: Johnson Mortimer.
Os Plesiossauros compunham o grupo de predadores marinhos mais bem-sucedidos e melhor distribuídos durante todo o Mesozóico. Tinham formas e tamanhos variados, que incluíam animais de pescoço longo com cabeça pequena ou de pescoço curtos com cabeça enorme, mas geralmente possuíam a cauda muito curta e as nadadeiras bem desenvolvidas.
O grupo surgiu no meio do Período Triássico e seguiu até o Maastrichiano (final do Período Cretáceo), onde enfrentou sua ext
inção, sem deixar descendentes de alguma forma. Há alguns que acreditam na existência de Plesiossauros vivendo isolados no Lago escocês Loch Ness. Até hoje, porém, não houve evidências concretas e toda aquela história não passa, na verdade, de mito.
inção, sem deixar descendentes de alguma forma. Há alguns que acreditam na existência de Plesiossauros vivendo isolados no Lago escocês Loch Ness. Até hoje, porém, não houve evidências concretas e toda aquela história não passa, na verdade, de mito.
Os Plesiossauros (Diapsida: Sauropterygia: Plesiosauria) eram compostos por dois grandes grupos: os Elasmossauros (com cabeças incrivelmente pequena e um pescoço muito longo. Figuras 3 e 4); e os Pliossauros (com pescoço curto e cabeças enormes).
Não há registro de fetos fósseis para o grupo, logo não é sabido se esses animais davam à luz aos seus filotes na agua, ou se desovavam na praia. A constituição robusta caixa toráxica foi justificada por alguns cientistas para defender a teoría de os Elasmossauros pudessem ir à praia, em terra firme. Entretanto, as nadadeiras compridas e os pescoços muito longos os tornariam muito desajeitados fora da água e eles se tornariam presas fáceis para dinosauros carnívoros ou crocodilianos. Atualmente, a grande maioria dos paleontólogos acreditam ser quase impossível que esses animais saíssem da água.

Figura 5: Reconstrução computadorizada de um Kronosaurus.

Figura 6 (autoria desconhecida): Reconstrução computadorizada de um Kronosaurus.
“O Liopleurodon ergue sua cabeça robusta vagarosamente e movimenta suas nadadeiras. À medida que ele avança, amonites agitam-se na agua e os peixes escondem-se nos corais em seu temor. Sua boca abre e atinge gravemente a porção do meio de um Ophthalmosaurus. A força de seu ataque carrega ambos sua cabeça e sua presa para fora da agua, onde, por um breve instante ele pausa antes de trazer ambos abaixo com uma força explosiva. Há sangue por todos os lados. Sua vítima more instantaneamente, seu corpo perfurado pelos longos dentes e suas costas quebradas. O Pliossauro ajeita sua presa na boca, mordendo e sacudindo-a repetidamente (…) Ele volta à superficie erguendo a garganta rosada e engolindo.”
(Haines)
Liopleurodon tratava-se de um Pliossauro. Colossais predadores, entre os maiores répteis carnívoros que já viveram.
As primeiras formas intermediárias entre os plesiossauros e os pliossauros surgiram no início do Jurássico, como os Macroplata longirostris, M. tenuiceps e possivelmente Eurycleidus arcuatus. Eram plessiossauros de pescoço mais curto e crânio ligeiramente mais robusto.
O gênero Pliosaurus é conhecido para o Jurássico Médio, e foi um dos primeiros do grupo a ter as características que os definem.
Ao Final do Jurássico e Cretáceo, diversas formas floresceram, incluso os colossais predadores Liopleurodon, Kronosaurus, Mareasaurus, Brachauchenius, Megalneusaurus e Peloneustes. Sua distribuição era ampla, incluindo as Américas, a Europa, a Ásia e a Oceania.
Em 1992, o Paleontólogo alemão Oliver Hampe descreveu um enorme Pliossauro proveniente da região de Boyacá, ao norte da Colômbia. Ele foi nomeado Kronosaurus boyacencis, embora suas costelas, demasiado robustas (diferente do encontrado em outros do grupo), pudessem ser peculiares o suficiente para que se levantasse a hipótese de um novo gênero para a América do Sul. Os Kronosaurus (figuras 5 e 6) estavam distribuídos desde a Austrália até a Colômbia.
Ao final do Cretáceo foram extintos, assim como tantas outras espécies marinhas.

Figura 7: Reconstrução computadorizada de Mosassauros.
Fonte: Johnson Mortimer.

Figura 8: Reconstrução de Mosassauro.
Fonte: Walter Colvin.
Os Mosassauros (Figuras 7 e 8) foram criaturas aparentadas aos lagartos varanídeos atuais. Eram extremamente bem adaptados à vida marinha: enormes predadores de corpo alongado, esguio, dentes triangulares afiados e uma comprida cauda que lhes dava propulsão para perseguir suas presas.
Esse grupo evoluiu rapidamente (em termos de escala geológica) durante meados do Período Cretáceo. Há 90 milhões de anos atrás, Mosassauros já habitavam diversas regiões do globo e estavam entre os animais marinhos mais bem-sucedidos daquele momento. Alguns pesquisadores sugeriram que os Mosassauros teriam gradualmente substituído o nicho ecológico dos Ictiossauros, que se extinguiram no início daquele Período. Entretanto, parece haver uma incompatibilidade em relação aos hábitos alimentares dos dois grupos para que sustentasse essa idéia.
Mosassauros, assim como diversos outros grupos, foram totalmente extintos durante o final do Cretáceo.
Bibliografia:
. Ellis, Richard. Sea Dragons: predators of the prehistoric oceans. University Press of Kansas, 2003.
. Motani, Ryosuke. The Evolution of Marine Reptiles. Evo Edu Outreach (2009) 2:224–235. Acesso livre em Springerlink.com, 2009















 William Roberto Nava
William Roberto Nava