Uma nova e interessante proposta para explicar as relações evolutivas dos dinossauros e principalmente, a origem dos dinossauros ornitísquios, grupo que inclui o Triceratops e os famosos dinossauros “bico de pato”, acaba de ser apresentada por dois pesquisadores brasileiros da Universidade Federal de Santa Maria (UFSM), RS, Rodrigo Temp Müller e Maurício Garcia. O estudo foi publicado na revista científica Biology Letters e agita ainda mais a discussão sobre as relações evolutivas dos dinossauros.

Não faz muito tempo que um estudo liderado por um pesquisador britânico chamado Matthew Baron, sacudiu a comunidade paleontológica e abriu espaço para novas discussões sobre uma questão fundamental no estudo de dinossauros: a relação evolutiva entre os grandes grupos desses animais.
Em 2017, Baron e colaboradores ressuscitaram uma antiga hipótese sobre a relação dos dinossauros, que propunha que o grupo que inclui os dinossauros carnívoros, como o Tyrannosaurus, era mais proximamente relacionado ao grupo dos dinossauros ornitísquios, aquele que reune Triceratops, Stegosaurus e os chamados “dinos bico de pato”. A hipótese de Baron e colegas veio com algumas novidades, mas já havia sido considerada no passado, tendo sido descartada por diversos estudos em detrimento da proposta mais clássica, que reúne dinossauros carnívoros e os grandes dinossauros “pescoçudos ” em um mesmo grupo chamado Saurischia.
Se quiser ler mais sobre essa história, clique aqui.
O agito causado por esse artigo foi tanto, que, na época, muito se falou na mídia popular, inclusive, como se tudo que sabíamos sobre dinossauros tivesse que ser reescrito. Pouco tempo passou, todavia, até a publicação de um outro estudo, rebatendo a proposta de Baron e colaboradores. Estudo esse, liderado por um pesquisador brasileiro, da USP de Ribeirão Preto, Prof. Max Langer. O estudo liderado por Langer apontava inconsistências no trabalho de Baron e concluía que a hipótese clássica da divisão dos dinossauros ainda se sustentava com prioridade, apesar de a proposta de Baron não poder ser totalmente descartada, já que era somente “um pouco menos provável que a hipótese tradicional”. Leia mais sobre isso aqui.
A história é longa e a disputa continuou em alguns estudos subsequentes, mas aonde eu quero chegar é que, toda essa discussão ressaltou novamente uma verdade muito incômoda para os estudiosos de dinossauros: nós conhecemos muito pouco sobre os primeiros ornitísquios e não conseguimos dizer com certeza como eles se relacionam com os outros dinossauros. E é aí, finalmente, que o trabalho recém-publicado pelos colegas da UFSM pode ajudar.
Os “caçadores de dinossauros” da UFSM
Rodrigo Temp Müller e Maurício Garcia são dois pesquisadores muito privilegiados, pois têm a honra de estar justamente no local que provavelmente foi o berço dos dinossauros, há aproximadamente 230 milhões de anos. Os fósseis de dinossauros mais antigos conhecidos no mundo são encontrados em rochas dessa idade na Argentina, no sul do Brasil e na porção sul do continente africano. Justamente por estarem trabalhando sobre essas rochas, esses pesquisadores têm acesso direto a fósseis fantástico, que contam a história da aurora desse grupo tão fascinante de animais.

Atualmente, Rodrigo T. Müller e Maurício Garcia, junto a outros pesquisadores, trabalham vinculados ao Centro de Apoio à Pesquisa Paleontológica (CAPPA-UFSM), em São João do Polêsine, RS, onde se dedicam a estudar a história dos primeiros dinossauros.
A nova hipótese de Müller e Garcia
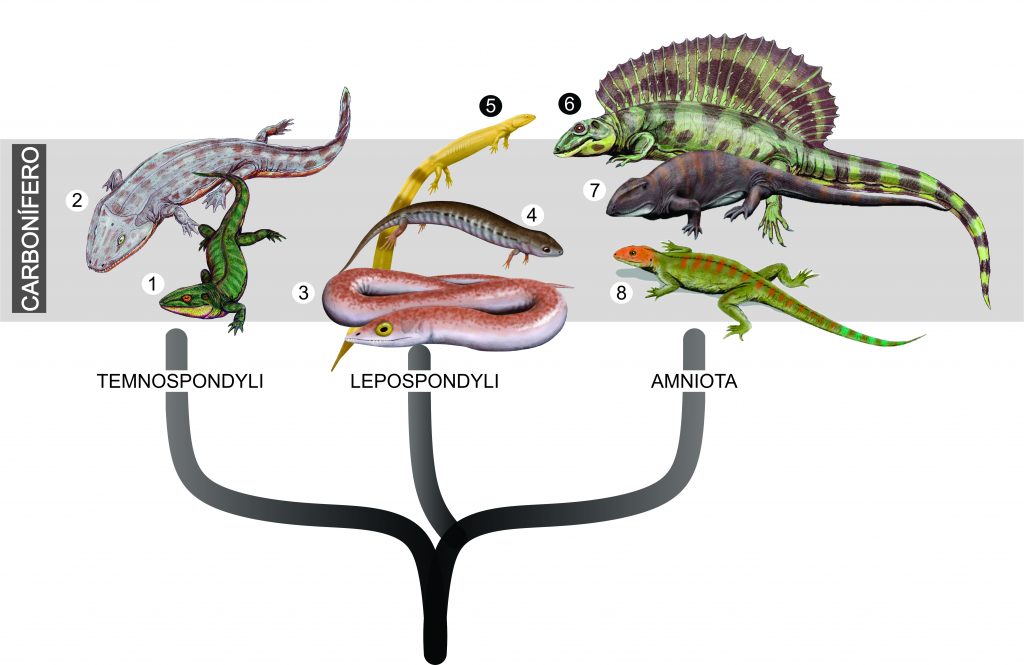
Verdade seja dita, a origem dos dinossauros ornitísquios é um grande mistério. Enquanto os fósseis mais antigos de dinossauros saurísquios datam de estratos do meio do Período Triássico, entre 245 e 230 milhões de anos, os fósseis mais antigos de ornitísquios são um pouco mais recentes, da transição Triássico-Jurássico, e têm aproximadamente 200 milhões de anos. Explicar esta ocorrência usando a hipótese tradicional sobre da relação dos dinossauros é um pouco constrangedor, pois implica na existência de “linhagens fantasmas” de ornitísquios, que teriam existido entre ‘pouco tempo depois da origem dos dinossauros’, por volta de 240-230 milhões de anos, até cerca de 206 milhões de anos atrás, idade do fóssil mais antigo de ornitísquio conhecido. Resumindo: ornitísquios necessariamente teriam que ter existido entre ~240 e 206 milhões de anos, mas não temos evidências dos fósseis deles.
A nova hipótese proposta por Müller e Garcia solucionaria esse problema de maneira muito elegante, com algo que sempre esteve bem debaixo do nariz dos pesquisadores: os “Silesauridae”.
As criaturas conhecidas como ‘silessaurídeos’ compõe um grupo de organismos extintos normalmente considerados como parentes muito próximos dos dinossauros. De fato, eles e os primeiros dinossauros são extremamente parecidos, tanto em forma, hábitos, quanto em tamanho (imagem).
Fósseis de ‘silessaurídeos’ são encontrados no Brasil, como é o caso de Sacisaurus agudoensis, um pequeno animal, com cerca de 1,5m de comprimento, que viveu há 225 milhões de anos onde hoje é o Rio Grande do Sul.

O que Müller e Garcia fizeram em seu estudo foi testar as relações evolutivas dos grandes grupos de dinossauros, incluindo uma ampla amostragem de silessaurídeos (até então tidos como grupo irmão de Dinosauria) e diversas espécies de dinossauros basais. Eles fizeram uma análise filogenética abrangente, que juntou, pela primeira, vez os dados de espécies há tempos conhecidas, com as espécies mais recentemente descritas. Os resultados encontrados por eles foram surpreendentes…
O que uma análise filogenética faz, basicamente, é testar as relações evolutivas dos organismos por meio da comparação de uma extensa matriz de dados sobre eles. Estes dados podem ser genéticos ou morfológicos, ou os dois, por exemplo. No caso dos fósseis, os pesquisadores normalmente usam dados morfológicos (da forma) para realizá-las, pois informações genéticas só ficam preservadas em materiais relativamente recentes (de até algumas centenas de milhares de anos). O produto de uma análise filogenética são um conjunto de “árvores evolutivas” possíveis, mostrando todas as relações prováveis entre os organismos analisados. Algumas árvores são estatisticamente mais plausíveis que outras e, normalmente são essas as consideradas mais atentamente pelos pesquisadores.
Uma incrível descoberta
O que Müller e Garcia recuperaram em suas análises foi algo diferente de tudo antes proposto e muito excitante para os estudiosos de dinossauros, já que tem o potencial de explicar várias questões sobre o início da história evolutiva do grupo. Em especial, o tal mistério das “linhagens fantasmas” de ornitísquios.
De acordo com os resultados do estudo, os chamados ‘silessaurídeos’ seriam, na verdade, uma série de espécies ordenadas sucessivamente na base de Ornithischia. Ou seja, linhagens que teriam gradualmente acumulado características típicas de ornitísquios ao longo de milhões de anos. Essa hipótese preencheria o intervalo evolutivo antes “assombrado” pela ideia de “linhagens fantasmas”.
Os “silessaurídeos” seriam os dinossauros ornitísquios basais “que faltavam”! De acordo com a hipótese recuperada na análise de Müller e Garcia (2020), ‘Silesauridae’ seriam um grupo parafilético, na base de Ornithiscia. Sob esta perspectiva, Sacisaurus voltaria a ser um dinossauro.

Outros autores já haviam apontado uma possível relação entre silessaurídeos e ornitísquios, mas o cenário evolutivo encontrado por Müller e Garcia é inédito. Os ornitísquios clássicos, segundo o novo estudo, teriam evoluído a partir de formas típicas de silessaurídeos por meio de mudanças graduais ao longo do tempo, partindo inclusive, de uma possível forma carnívora. O que assemelharia o cenário evolutivo dos ornitísquios ao dos saurísquios sauropodomorfos (veja este vídeo aqui para entender).
Todos os ornitísquios conhecidos até o momento apresentam características que os vinculam a uma dieta herbívora, todavia os mais antigos ‘silessaurídeos’ tinham dentes pontiagudos e afiados, possivelmente adaptados para uma dieta carnívora.
Os pesquisadores responsáveis pela pesquisa reconhecem que ainda é cedo para que a questão relacionada a origem dos dinossauros ornitísquios possa ser considerada totalmente respondida. A nova hipótese de Müller e Garcia agora deverá continuar sendo testada à medida que novos fósseis foram descobertos e novas interpretações realizadas.
Nós aqui também estamos de olho.


Se você se interessou pelo estudo, você pode ler ele na íntegra AQUI.
Leia também a postagem no blog do parceiro ‘Coelho Pré-Cambriano’: AQUI.
Referência:
Müller, R.T. & Garcia, M. 2020. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biology Letters, https://doi.org/10.1098/rsbl.2020.0417