


Por que ATP!?
![]()
A maioria das pessoas que sabe um pouquinho de bioquímica aprende que o ATP é a “moeda energética” da célula. Maravilha. Mas aposto que se você perguntar o porquê disso, a resposta-padrão vai ser: “É porque é uma molécula muito energética, presente em reações de catálise e regulação de diversas enzimas na célula e blá-blá-blá…”. Lindo. Mas existem muitas moléculas muito energéticas no universo, e argumentar que o ATP é a “moeda energética” simplesmente porque as pessoas descobriram que na natureza ele está presente em várias reações enzimáticas acaba sendo um jeito bastante sofisticado de dizer “porque sim, zequinha!”.
[youtube_sc url=”https://www.youtube.com/watch?v=8Wldlug58Kc”]
Como construir um regulador enzimático e transportador energético na célula!?
Seguindo o espírito de engenheirar o metabolismo, como poderíamos, conhecendo as características físico-químicas dos elementos e compostos químicos, construir algo que seja como o ATP: uma coisa que regule de maneira sistêmica as atividades enzimáticas da célula, armazene energia e que seja fácil de se armazenar e ao mesmo de se usar!?
Bem, vamos pensar em energia como dinheiro: digamos que você seja rico, muito rico. Com muito dinheiro, a probabilidade de que você seja roubado é bem maior. Para que você mantenha sua riqueza é preciso que ela seja armazenada de um jeito “estável” (doce ilusão, mas considere isso como verdade, ok?) e por isso você coloca o seu dinheiro no banco. Você precisa proteger seu dinheiro e ainda acessá-lo facilmente – colocar tudo num baú e enterrar numa ilha desconhecida não vai ajudar na funcionalidade do seu dinheiro.
O mesmo funciona para a nossa moeda energética: ela não pode ser um composto cuja energia possa ser facilmente “roubada” por qualquer espécie química, mas também não pode ser pouco reativa a ponto de não conseguir regular a atividade enzimática e a transmissão energética. É preciso ter o controle desse “dinheiro” para gastá-lo facilmente onde você precisa: na manutenção da vida celular.
Então temos três coisas principais para a nossa molécula-moeda-energética: ela precisa ser estável o suficiente para que a energia não se “esvaia” facilmente, não tão estável a ponto de ser difícil usá-la e ainda reagir facilmente com proteínas.
Candidatos
Sabemos experimentalmente que o fosfato do ATP é a molécula-moeda. Mas porque só ela? Será que não tem nenhuma outra molécula que poderia fazer isso? Não poderia existir nenhuma outra coisa (ou coisas) alternativa que pudesse substituir o ATP ou que possa estar fazendo a mesma coisa?
Para encontrar um substituto (ou equivalente) para o ATP é preciso explorar análogos estruturais e energéticos dessa molécula. Selecionei dois possíveis candidatos:
TriSulfonatos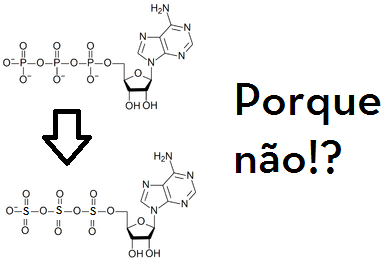
Da mesma forma que o ATP é um TriFosfato, porque ele não poderia ser um trisulfonato? A estrutura é bem similar e as energias das ligações éter (a ligação S – O – S) são bem parecidas também (veja). Além disso, sabe-se que há resíduos de aminoácidos que sofrem sulfonização.
Arsênico
Do mesmo jeito que teorizam as diferentes maneiras da vida ter surgido no universo, porque não 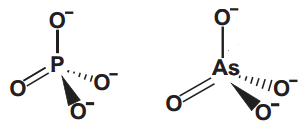 poderíamos ter um “ATP de Arsênico”!? O Arsênico fica logo abaixo do fósforo na tabela periódica e por isso faz o mesmo número de ligações, além de formar arsenato (e com isso diarsenatos e triarsenatos), de estrutura bem parecida com a do fosfato.
poderíamos ter um “ATP de Arsênico”!? O Arsênico fica logo abaixo do fósforo na tabela periódica e por isso faz o mesmo número de ligações, além de formar arsenato (e com isso diarsenatos e triarsenatos), de estrutura bem parecida com a do fosfato.
Que o melhor ganhe
As reações que o ATP faz são reações de hidrólise: um fosfato tem sua ligação éter (O – P) quebrada resultando na fosforilação de alguma coisa. Essa ligação é de alta energia (em termos bioquímicos) e é espontânea (delta G negativo). Reações não termodinamicamente espontâneas que passam por uma etapa de fosforilação acabam se tornando espontâneas na soma geral da energia livre, como por exemplo na fosforilação da glicose:
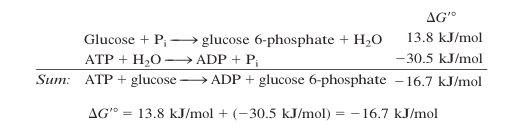
(Imagem modificada da referência [6])
Apesar da reação de hidrólise ser espontânea, a fosforilação tem uma velocidade de reação suficientemente lenta para garantir a estabilidade que a “moeda energética” precisa para realizar reações úteis (não que hidrolisar e simplesmente gerar calor seja de todo inútil…).
Se eu conseguir mostrar que o arsenato ou o sulfonato são melhores em termos energéticos e cinéticos, um mundo novo de possibilidades bioquímicas se abrirá!
Pesquisei alguns dados na internet e montei a seguinte tabela:
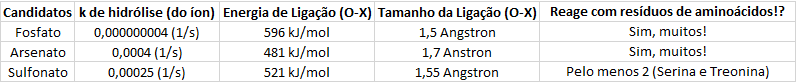
(Dados retirados das refs [1], [2], [3] e [4])
O quão maior for a constante de hidrólise (k) da reação, mais rápida ela será. Com esses dados dá para ver claramente que o fosfato é um “banco energético” bem mais confiável que os arsenato e o sulfato, sendo cerca de 100000 vezes mais estável que estes dois últimos – mantendo sua facilidade de “uso” energético com o delta G de hidrólise negativo. O fosfato é, além disso, bem mais “rico” que os seus concorrentes.
O sulfato, apesar de ser um candidato tentador, é bem mais difícil se “ser um ATP” como o arsenato pode ser. Não consegui encontrar nenhuma molécula como a proposta na imagem anterior, apenas dissulfonatos – e ainda não ligados a uma adenosina! Mesmo que houvesse um “ATP de enxofre”, seria difícil que ele assumisse o papel de controle das atividades enzimáticas porque ele não reage com resíduos de aminoácidos tão bem como o arsenato e o fosfato.
O arsenato já ganhou espaço na mídia a algum tempo atrás com o suposto encontro de uma “vida baseada em arsênico” na terra. Existem de fato várias enzimas que se ligam a arsenato e ele consegue substituir o fosfato em muitos casos, mas causando efeitos tóxicos na célula ao competir com o fosfato para a fosforilação de moléculas durante o catabolismo. Mesmo que houvesse um organismo em que a toxicidade fosse tolerada, a eficiência do processo de “arsenização” seria pífia: muitas moléculas apenas iriam hidrolisar espontaneamente de maneira “inútil”, liberando calor – além de tudo isso, eu nem preciso argumentar a questão a abundância do Arsênico no meio ambiente!
Não é porque sim!
Enfim, agora estou convencido do porquê o ATP é como é: o fósforo e as ligações éter o fazem um composto único, com propriedades flexíveis e ideais para ser o transmissor de energia de compostos mais energéticos do metabolismo até os menos energéticos, fazendo reações termodinamicamente não-espontâneas acontecerem e regulando de maneira sistêmica todo o conjunto de milhares de enzimas das células que dependem da fosforilação para alternarem entre seus estados ativos e inativos.
Portanto, da próxima vez que você não encontrar a resposta que queria, é melhor tentar construir o que você não entende – afinal, qual seria melhor jeito de entender alguma coisa no final das contas!?
Referências
[1] Rosen, B., Ajees, A., & McDermott, T. (2011). Life and death with arsenic BioEssays, 33 (5), 350-357 DOI: 10.1002/bies.201100012
[5] Tabelas de Entalpias de Ligação
[6] Nelson, David L., Albert L. Lehninger, and Michael M. Cox. Lehninger principles of biochemistry. Macmillan, 2008.