Durante do Cretáceo Superior, há cerca de 80 milhões de anos atrás, a região hoje correspondente ao noroeste do estado de São Paulo e Triângulo Mineiro em Minas Gerais, abrigava uma rica e diversificada fauna de crocodiliformes terrestres (parentes distantes dos crocodilos e jacarés atuais), que prosperava em meio aos gigantes dinossauros.
Recentemente uma nova espécie de crocodiliforme desse período foi descoberta em uma cidade do interior de São Paulo. Descrita por pesquisadores brasileiros, o novo animal, com o crânio extraordinariamente bem preservado, ganhou um nome de dar medo: Gondwanasuchus scabrosus. Quer entender o por quê desse estranho nome de batismo? Vamos primeiro conhecer um pouco mais sobre esse animal:
Arte de Rodolfo Nogueira.
– Texto por Thiago Marinho –
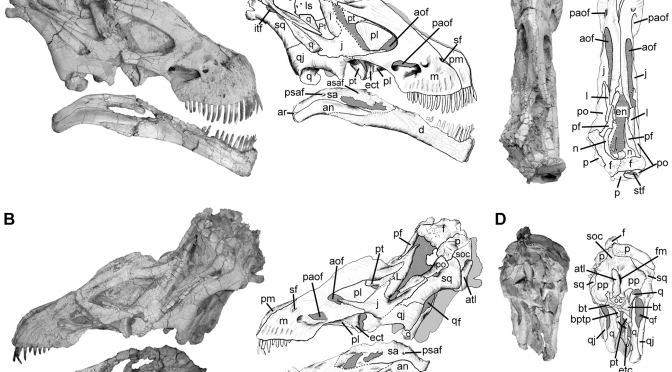
O mais novo representante dos crocodiliformes terrestres do Cretáceo do Brasil, Gondwanasuchus scabrosus, é um pequeno predador da Família Baurusuchidae, composta por importantes predadores e carniceiros que poderiam até mesmo competir por presas com pequenos dinossauros. Essa nova espécie foi descrita com base em um crânio parcialmente completo e muito bem preservado, proveniente de rochas da Formação Adamantina do município de General Salgado, noroeste do estado de São Paulo.Gondwanasuchus não passaria de 1,30 m de comprimento, mas o que esses animais não tinham em tamanho, tinham em adaptações que os tornavam eficientes predadores.
O nome do gênero, Gondwanasuchus, faz alusão a distribuição da família dos baurussuquídeos, restrita a regiões do antigo supercontinente Gondwana (que durante o Cretáceo agrupava a América do Sul, África, Madagascar, Índia, Oceania e Antártica) e, suchus, que significa crocodilo. O nome que define a espécie, scabrosus, é uma palavra em Latim que significa “tinhoso”, um apelido dado pelos pesquisadores que descreveram a espécie, devido à aparência “mal-encarada” do animal.
O fóssil de Gondwanasuchus scabrosus é representado por um crânio parcialmente completo, que foi encontrado em 2008 em associação a um grande indivíduo de Baurusuchus salgadoensis, um crocodiliforme também da família dos baurussuquídeos. Gondwanasuchus scabrosus convivia não só com outros baurussuquídeos, mas também com crocodiliformes herbívoros da família dos esfagessaurídeos. A presença de esfagessaurídeos e o fato de os depósitos da Formação Adamantina no município de General Salgado serem basicamente compostos por paleossolos (solos que foram preservados no registro geológico), sugerem que pelo menos algumas partes do habitat de Gondwanasuchus eram compostas por áreas com vegetação arbustiva e arbórea.
Crânio peculiar:
O crânio de Gondwanasuchus é altamente comprimido lateralmente, como o de muitos dinossauros carnívoros – bastante diferente dos crocodilos atuais! Suas narinas eram posicionadas lateralmente na região anterior do focinho e o animal possuía grandes órbitas oculares voltadas para frente.
Dentes modificados:
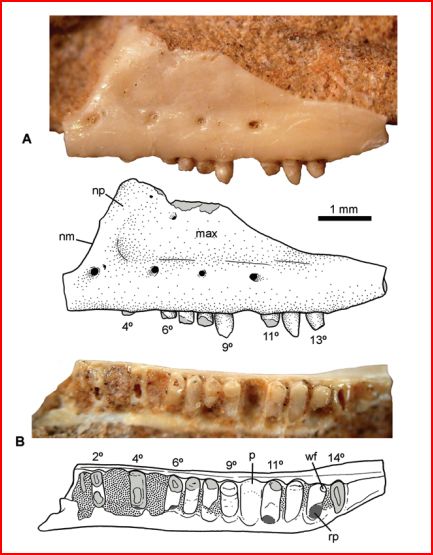
Os dentes posteriores de G. scabrosus são altamente comprimidos e com bordas serrilhadas, como os dentes de alguns dinossauros carnívoros. Outra peculiaridade da dentição desses animais é a presença de profundas estrias que percorrem os dentes da base para o topo, possivelmente garantindo uma maior resistência a quebra durante o processo de caça e alimentação.
Visão especializada:
Os olhos de Gondwanasuchus scabrosus eram destacadamente voltados para frente, diferentemente do observado na maioria dos outros crocodiliformes, que possuem os olhos orientados lateralmente. Essa característica permitia que esses animais tivessem visão binocular, ou seja, eles poderiam enxergar tridimensionalmente, o que seria muito útil para uma melhor avaliação da distância dos objetos observados e melhor precisão de seus ataques.
Quer mais detalhes? Clique no infográfico para ampliar!
Clique para ampliar
Interessado em mais informações sobre esse animal?? Escreva pra gente (colecionadoresdeossos@gmail.com)!! Thiago da Silva Marinho, o primeiro autor do artigo, é membro aqui do Colecionadores de Ossos!!
Thiago da Silva Marinho Biólogo pela Universidade Federal de Uberlândia (UFU), Mestre e Doutor em Geologia pela Universidade Federal do Rio de Janeiro (UFRJ), hoje é professor efetivo da Universidade Federal do Triângulo Mineiro (UFTM).
Desenvolve estudos com ênfase em arcossauros mesozóicos, especialmente crocodyliformes e dinossauros. Clique aqui para ver o Currículo Lattes.
MARINHO, T. S. et al. Gondwanasuchus scabrosus gen. et sp. nov., a new terrestrial predatory crocodyliform (Mesoeucrocodylia: Baurusuchidae) from the Late Cretaceous Bauru Basin of Brazil. Cretaceous Research. 2013 (on-line).
O bicho é meu e eu coloco o nome que eu quiser nele!!!
Pintossauro, Gasossauro, Dinheirossauro, Fodonyx, Bambiraptor…? A lista é longa! Um mais estranho que o outro! Mas porque eles foram batizados assim?
Veja aqui uma amostra dos nomes mais estranhos da Paleontologia!
Ai Ai Estou Morrendo Seu Idiota da Silva Sauro
Quem não lembra do episódio da Família Dinossauro em que Baby é levado para o grande sábio afim de receber um nome, mas algo inesperado acontece? Ele acaba recebendo um nome meio que, digamos… incomum: “Ai Ai Estou Morrendo Seu Idiota da Silva Sauro”.
(se você nunca assistiu, veja o episódio AQUI 🙂 ).
Isso nos leva a pensar: Afinal, como são escolhidos os nomes dos dinossauros??
Certamente não é como no episódio da família dinossauro (ver post anterior), mas ainda assim parecem surgir alguns resultados meio… incomuns.
Neste post vamos reunir alguns dos nomes mais inusitados já escolhidos por paleontólogos.
Não são só os dinossauros que sofrem, mas toda ‘sorte’ de criatura extinta…
————
Os 10 mais!
And the Oscar goes to:
1) PINTOSAURUS: gênero basal de procolofonídeo. Procolofoquê?Procolofonídeo! Procolofonídeos são pararépteis que lembram muito um “tipo robusto de lagarto”, mas na verdade não tem nada a ver com eles (Veja AQUI e AQUI para saber mais). Pintosaurus foi descrito em 2004, encontrado em rochas de idade Permo-triássicas do Uruguai. Pinto- foi escolhido para homenagear o Dr. Iraja Damiani Pinto, paleontólogo gaúcho que contribuiu substancialmente para paleontologia sulamericana. (…)
2) CHUPACABRACHELYS: Trata-se de uma tartaruga do período Cretáceo, que foi encontrada no Texas, USA. Não tem muito a dizer… o nome realmente foi em homenagem ao “chupa-cabra”…
3) GASOSAURUS CONSTRUCTUS: Dinossauro terópode chinês de médio porte. O nome foi escolhido para homenagear uma empresa de combustível. O dinossauro foi encontrado durante sua construção, por isso o nome da espécie é ‘constructus‘.
4) DINHEIROSAURUS LOURINHANENSIS: Uma espécie de dinossauro saurópode gigante, aparentado ao Diplodocus. Ele foi encontrado na região da Praia de Porto Dinheiro, concelho de Lourinhã, em Portugal. O nome deriva do local aonde ele foi encontrado. Tinha que ser um dinossauro português..
5) BAMBIRAPTOR: Sim, esse é um dinossauro que foi nomeado em homenagem ao Bambi. Isso mesmo, aquele personagem da Disney (…!!). Trata-se de um pequeno dinossauro carnívoro, com menos de 1m de comprimento. O nome foi escolhido porque aparentemente o espécime encontrado era um juvenil.
6) FODONYX: Rincossauro do Triássico Médio da Inglaterra. O nome significa “garra escavadora”. Em latim “fodere“=escavar e “onyx“=garra. “Tchau! vou ‘fodere’ dinossauros!”
7) MINOTAURASAURUS: Tipo de dinossauro anquilossaurídeo proveniente da Ásia. Foi descrito em 2008. O nome significa “Homem-touro-lagarto”.
8) GOJIRASAURUS: Gênero dúbio de dinossauro terópode encontrado em rochas triássicas do Novo México, EUA. O nome foi em homenagem ao monstro mitológico japonês. Gojira…. Gojira….Gojira…
Incisivosaurus – não é só o nome que é feio…
9) INCISIVOSAURUS: Um pequeno dinossauro terópode da China, provavelmente de hábito herbívoro. Sua dentição peculiar, com dentes proeminentes como os e um roedor, foi o que lhe rendeu o nome estranho.
10) PIKAIA: Representante basal do grupo dos cordados. O nome dessa criaturinha Cambriana encontrada em Burgess Shale significa “da Pika”. Pika é um tipo de pequeno mamífero aparentado dos coelhos, comumente encontrado na região aonde os fósseis de Pikaia foram descobertos. (Eu ri!)
——–
No mínimo exóticos:
– Enigmosaurus: Grande dinossauro terópode herbívoro (Therezinosauróide) do Cretáceo da Mongólia. Quando os seus restos foram encontrados, a anatomia não usual da pélvis do bicho deixou os seus descobridores tão confusos, que lhes pareceu um verdadeiro enigma. Assim sendo, resolveram nomeá-lo dessa forma.
– Camelotia: Gênero de um dinossauro prossaurópode do Triássico Inglaterra. Seu nome significa “de Camelot”. Camelot trata-se do lugar lendário na Grã-Bretanha, que teria abrigado o castelo e a corte do Rei Arthur.
– Erectopus: Dinossauro terópode allossauróide do Cretáceo da França. Descoberto no final do Século XIX, o seu nome significa: “Erecto=em pé”, “Pous=Pé”.Hm…
– Minmi: Dinossauro australiano da infraordem Ankylosauria. O nome é devido à Minmi Crossing, o lugar onde seus fósseis foram descobertos.
– Hallucigenia: Gênero de invertebrado fóssil do Período Cambriano. O nome é devido a sua forma bizarra, que aos olhos dos descobridores mais parecia uma alucinação. Já viu né!!
– Drinker: Pequeno dinossauro hypsilofodontídeo do Jurássico da América do Norte. O nome, traduzido do inglês significa “bebedor”, mas essa não foi a intenção, ele foi proposto para homenagear o renomado paleontólogo Edward Drinker Cope.
– Gargoyleosaurus: Um dos mais antigos anquilossauros já descobertos. Gargoyleasaurus foi encontrado em Wyoming, em rochas de idade Jurássica. Seu nome significa “lagarto gárgula”.
– Xixiasaurus: Dinossauro terópode troodontídeo descrito em 2010. Seus restos foram encontrados na região administrativa de Xixia, na província de Henan, China.
– Pawpawsaurus: Gênero de dinossauro da família Nodosauridae, da infraordem Ankylosauria. Seus restos foram encontrados na Formação Paw Paw, Texas, USA. Eu não consigo falar sem rir! – Pedopenna: Dinossauro manirraptor do Jurássico da China. Pedopenna significa “pena no pé”. Este dinossauro recebeu esse nome por apresentar evidências da existência de longas penas inseridas ao longo de seus metatarsos. – Ozraptor: Dinossauro terópode encontrado na Austrália, descrito em 1998. “Oz” faz referência ao apelido dado aos australianos,“Ozzies”. – Borogovia: O nome deste dinossauro carnívoro troodontídeo é derivado dos “borogoves”, criaturas de um poema de Lewis Carroll, Jabberwocky, parte da obra “Alice no país das Maravilhas”. – Appalachiosaurus: Gênero de dinossauro terópode tiranossauróide do Cretáceo da América do Norte. O nome faz referência a região estadunidense conhecida como Appalachia, onde o fóssil do animal foi encontrado.
– Petrobrassaurus puestohernandezi: Dinossauro saurópode argentino. Foi descrito em 2011. O nome do gênero realmente é devido a companhia de petróleo brasileira, Petrobrás, e o nome específico se refere a “Puesto Hernandez”, um dos centros de extração da companhia, aonde o dinossauro foi encontrado. Ahhh, invejosos! Queridos!!
– Atlascopcosaurus: Este dinossauro australiano recebeu seu nome em homenagem a companhia Atlas Copco. Esta companhia forneceu o equipamento para a expedição paleontológica que resultou na descoberta do novo dinossauro. Capessaurus, Fapespsaurus, Cnpqsaurus, vamo lá, galera!!
– Qantassaurus: Este dinossauro hypsilofodontídeo foi nomeado para homenagear a empresa aérea australiana, Qantas, que ajudou no transporte dos fósseis. Conclusão: australiano não sabe dar nome pra dinossauro!!
– Panamericansaurus: Outro gênero de dinossauro saurópode da Patagônia argentina. Foi descrito em 2010. O nome foi para homenagear a companhia petrolífera Pan American Energy, que deu apoio financeiro às pesquisas.
– Argentinosaurus – Ah…Esse só pra sacanear mesmo. Bonito nome.
—————-
Homenagens curiosas:
Utahraptor spielbergi – Esse dinossauro raptor foi descoberto na semana de estréia do filme Jurassic Park. O nome da espécie foi escolhido para homenagear Stephen Spielberg, diretor de “Jurassic Park”. Posteriormente re-descrito, o nome da espécie mudou para ostrommaysorum. Depois de ganhar milhões com Jurassic Park, acho que S. Spielberg não deve ter ficado chateado….
Tianchisaurus nedegoapeferima(Informalmente chamado de Jurassosaurus) – Tendo doado dinheiro para pesquisa de dinossauros na China, Stephen Spielberg sugeriu para esse dinossauro o nome de Jurassosaurus – em razão do lançamento do filme Jurassic Park em 1993. O nome esdrúxulo só pegou na informalidade. Porém ainda assim, o nome da espécie (nedegoapeferima) homenageia Jurassic Park: ele é formado pelas letras iniciais dos sobrenomes dos principais atores/atrizes que participaram do filme: Sam Neil, Laura Dern, Jeff Goldblum, Richard Attenborough, Bob Peck, MartinFerrero, Ariana Richards e Joseph Mazzello.
Dracorex hogwarsia– O coitado desse bicho foi batizado como um tributo a obra de J.K. Rowling, “Harry Potter“. – hogwarsia faz alusão a ‘Hogwarts’, a escola de magia.
Mimatuta morgoth – O Professor da Universidade de Chigado, Leigh Van Valen, nomeou uma série de mamíferos paleocênicos com base em personagens da série de livros “Senhor dos Anéis”. Morgoth foi em homenagem ao “The Dark Lord”, mas além dele ainda temos:
Alletodon mellon – mellon é a palavra élfica para “amigo” e a senha para a entrada nas minas de Moria.
Mithrandir onostus – Mithrandir sendo outro nome para Gandalf.
Oxyprimus galadrielae – Em homenagem a Lady Galabriel.
Protungulatum gorgun – ‘gorguns’ são os orcs.
Bom gosto esse cara.
Masiakasaurus knopfleri – Esse pequeno dinossauro predador recebeu seu nome em homenagem a Mark Knopfler, guitarrista da banda Dire Straits. De acordo com Scott Sampson, seu descobridor, o time resolveu batizar assim o dinossauro depois de escutar Dire Straits durante a escavação. De acordo com eles, novos dinossauros só eram encontrados quando esse som estava no rádio. Já tentamos essa tática, mas pra gente só funciona com “Hotel California” do Eagles… #sarcasmo
Aegrotocatellus jaggeri– Essa espécie de trilobita foi nomeada realmente a fim de homenagear estrelas do rock! O nome da espécie (jaggeri) foi um tributo a Mick Jagger, vocalista do Rolling Stones, enquanto que o nome do gênero “Aegrotocatellus” significa em latim “Sick Puppie” (ver a banda de rock alternativo australiana ‘Sick Puppies‘).
Jagger e seu trilobita
Um dos caras que ajudou a descrever esse trilobita (Greg Edgecombe), não satisfeito, nomeou uma outra série desses artrópodes primitivos com nomes de integrantes de bandas! Ele homenageou Sex Pistols (Arcticalymene viciousi, A. Rotteni, A. jonesi, A. cooki, A. matlocki), Ramones (Mackenziurus johnnyi, M. joeyi, M. deedeei, M. ceejayi), John Lennon, Ringo Star e Simon & Garfunkel (Avalanchurus lennoni, A. starri, A. simoni, A. Garfunkeli).
Norasaphus monroae–Esse trilobita foi uma homenagem de Richard Fortey, paleontólogo inglês, a Marlin Monroe.
– Piatzinigkosaurus– WTF? Isso foi sacanagem dos argentinos.
– Huehuecanauhtlus tiquichensis– uheuheueheuh!!
– Nqwebasaurus– Primeiro dinossauro com o som de “CLICK” em seu nome. Pronuncia-se: N – (som de click com a língua) – KWE – BA – SAU – RUS. Nqweba é o nome do lugar aonde o dinossauro foi encontrado, na África do Sul. Trata-se de uma palavra na língua da tribo Bantu.
– Jinfengopteryx– China 1
– Jingshanosaurus– China 2
– Szechuanosaurus– China 3
– Jinzhousaurus– China 4. Os nomes chineses sempre são os mais impossíveis!
– Phuwiangosaurus– Tailândia.
– Bruhathkayosaurus– Índia. Correção: Os nomes asiáticos são sempre os mais impossíveis.
– Parapropalaehoplophorus– fale-3-vezes-rápido!
–> Agora-fale-tudo-junto!!!!
.
—————-
“Éééé do brasiiilll!!!”
Como se já não bastasse oBaurusuchus e lanches salgadoensis (Veja o post anterior), ainda temos uma série de crocodilos tupiniquins com nomes estranhos:
– Morrinhosuchus – Ganhou seu nome em homenagem a um morro (!) que fica próximo ao local de coleta do fóssil, o “Morrinho de Santa Luzia”.
– Barreirosuchus – Mais uma alusão ao local de coleta, o bairro de Barreiros, em Monte Alto, SP;
– Pepesuchus – O nome foi uma homenagem ao Prof. José Martin Suárez (conhecido pelos colegas como Pepe).
Fora os dinossauros:
– Oxalaia quilombensis– O gênero é uma referência a divindade africana ‘Oxalá’ e a espécie refere-se aos alojamentos quilombolas da Ilha do Cajual (local onde o fóssil foi encontrado).
– Irritator challengeri – Dinossauro brasileiro nomeado por pesquisadores estrangeiros (David Martill e colegas) que ficaram “extremamente irritados” devido a “restauração” feita pelo seu coletor amador. Buscando fazer o fóssil parecer mais completo e valioso, o coletor clandestino acabou obscurecendo a real natureza do animal e dificultando o trabalho dos pesquisadores.Isso que dá comprar fóssil ilegalmente Dr. Martill!!!O nome da espécie foi uma homenagem ao Prof. Challenger, personagem do livro “O Mundo Perdido” de Arthus Conan Doyle.
* Agradecimentos aos colegas paleontólogos que contribuíram ajudando (pelo facebook!) a reunir os nomes mais bizarros de seus respectivos campos do conhecimento!
Ilustração acima: Danielle Byerley, Florida Museum of Natural History
A Formação Cerrejo, no Nordeste Colombiano, estimada como do Paleoceno Médio-Tardio – baseado em estudos de isótopos de carbono e palinomorfos – apresenta o primeiro bom registro de ecossistemas continentais terrestres para essa brecha no tempo na América do Sul.
As descobertas mais recentes, feitas numa área de exploração de carvão, indicam não só a mais antiga evidência macrofóssil de florestas neotropicais, como também, a presença da maior serpente fóssil já descoberta, Titanoboa cerrejonensis. Além disso, há registros de cágados gigantes, peixes dipnóicos e elopomorfos e de um raro Crocodiliforme Dyrossaurídeo de focinho curto, conhecido como Cerrejonisuchus improcerus.
Num trabalho recém-publicado de Alexander Hastings, Jonathan Bloch e Carlos Jaramillo, um novo dyrossaurídeo de focinho longo, Acherontisuchus guajiraensis foi descrito para a região. O material fóssil havia sido coletado em 1994, mas ficou retido na mina de carvão até 2004, quando foi levado a Universidade da Flórida para estudos. O que chama atenção, é o tamanho do animal.
Há 60 milhões de anos atrás, Acherontisuchus guajiraensis, de 9 m de comprimento, coexistiu com a gigantesca Titanoboa cerrejonensis em um sistema de rios semelhante ao atual Amazonas, atravessando uma as primeiras florestas tropicais úmidas do mundo, até desembocar no que seria um dia o Mar do Caribe.
O focinho longo do Acherontisuchus deve ter sido utilizado para capturar peixes e os autores ainda sugerem que o animal deveria ter competido com Titanoboa por alimento.
As análises nos fósseis descritos no artigo mencionado ecaixam Acherontisuchus em um grupo de Crocodiliformes chamado de Dyrossaurídeos, animais que usualmente habitavam ambientes marinhos e fluviais costeiros.
Vale à pena checar o artigo também porque os autores realizam uma nova análise cladística para os Dyrosauridae, utilizando 82 caracteres do crânio e mandíbula para encaixar Acherontisuchus guajiraensis em sua posição dentro do grupo. Eles incluem dyrossaurídeos do Novo Mundo e do Velho Mundo como Hyposaurus rogersii, Congosaurus bequaerti, Atlantosuchus coupatezi, Guarinisuchus munizi, Rhabdognathus keiniensis e Rhabdognathus aslerensis.
Os resultados são consistentes com relação a origem africana dos Dyrosauridae, com dispersões para o Novo Mundo durante o Cretáceo Tardio, passando por uma transição de habitats marinhos em taxa ancestrais, para ambientes fluviais em taxa mais derivados.
Bibliografia:
. HASTINGS, A.K., BLOCH, J. I., JARAMILLO,.C.A. 2011. A NEW LONGIROSTRINE DYROSAURID (CROCODYLOMORPHA, MESOEUCROCODYLIA) FROM THE PALEOCENE OF NORTH-EASTERN COLOMBIA: BIOGEOGRAPHIC AND BEHAVIOURAL IMPLICATIONS FOR NEW-WORLD DYROSAURIDAE. [Palaeontology, Vol. 54, Part 5, 2011, pp. 1095–1116]
Neste último mês, novas descobertas sobre o Cretáceo brasileiro vieram afirmar que neste período os Crocodyliformes realmente dominavam nosso país. Enquanto os dinossauros reinavam com soberania na maioria dos ecossistemas terrestres, aqui no território tupiniquim os crocodyliformes se diversificavam e ocupavam os mais diversos nichos. Desde animais com cerca de 4 metros de comprimento, ágeis e carnívoros, como o Baurusuchus, até pequenos onívoros, como o Mariliasuchus e o Adamantinasuchus. Haviam espécies escavadoras, oportunistas e até mesmo consumidoras de plantas e raízes.
Estas descobertas têm elucidado como teriam sido as relações ecológicas pretéritas do paleoambiente hoje representados pelas rochas do Grupo Bauru. As rochas do Grupo Bauru são de idade Neocretácica (Final do período Cretáceo, do Cenomaniano ao Maastrichiano, 99-65 milhões de anos atrás) e foram depositadas em um contexto continental fluvial e lacustre de clima quente e árido. Elas estão bem distribuídas nos estados de São Paulo e Minhas Gerais, mas também são encontradas no Paraná e Goiás, e até mesmo no estado do Mato Grosso. Estas rochas representam um antigo ecossistema que possuía uma diversificada fauna de vertebrados, incluindo dinossauros saurópodes (titanossauros) e terópodes (abelissauros, carcharodontossauros e maniraptores, incluindo as aves), lagartos, cobras, quelônios, anfíbios, pequenos mamíferos, mas principalmente crocodyliformes. Estes últimos englobavam até seis distintos grupos: os notossuquídeos, os sphagessaurídeos, candidodontídeos, peirossaurídeos, trematocampsídeos e baurussuquídeos.
Ao que se deve esta incrível diversidade de crocodilomorfos? O que pode ter favorecido o desenvolvimente deste grupo de arcossauros neste particular contexto pretérito? Os pesquisadores ainda procuram uma resposta. A solução para o enigma pode estar envolvida com o fato deste local pretérito ter sido geograficamente isolado e ter produzido uma situação ecológica e ambiental única, que favoreceu estes animais. Teriam eles ocupado o nicho até mesmo de dinossauros? Competido com eles? Ou o nicho de mamíferos, tão raros neste registro por algum propósito? A continuidade dos estudos vai ajudar a resposder estas perguntas.
Campinasuchus, o novo crocodyliforme do Cretáceo brasileiro
Fantástica reconstituição artística de Campinasuchus em vida por Rodolfo Nogueira.
Campinasuchus é um novo gênero de Baurusuchidae descrito com base em alguns crânios parciais e esqueletos encontrados na região de Campina Verde, MG, contexto da Formação Adamantina, Grupo Bauru, Bacia Bauru.
Os Baurusuchidae incluem crocodyliformes com crânios lateralmente comprimidos e gracilmente alongados. São conhecidos para o Cretáceo Tardio do Brasil, Argentina e Paquistão. Todos os membros podem ser considerados de médio e grande porte, cursoriais (caminhavam ativamente sem encostar a barriga no chão, com os membros posicionados mais verticalmente) e predadores. Outras espécies de Baurusuchidae incluem: Baurusuchus pachecoi, Baurusuchus salgadoensis, Baurusuchus albertoi e Stratiosuchus maxhechti do Brasil, além de Cynodontosuchus e Wargosuchus da Argentina.
Campinasuchus se diferencia dos outos Baurusuchidae por possuir um focinho mais curto e afilado, uma dentição diferenciada e peculiaridades no seu osso palatal (céu da boca).
A sua presença reforça a idéia de que a aridez, ou possivelmente um regime específico de sazonalidade (alternância de períodos quentes e secos com períodos de alta pluviosidade), dirigiram a diversificação dos crocodyliformes terrestres neste ecossistema peculiar do Cretáceo Tardio brasileiro.
O trabalho foi publicado por Ismar de Souza e Carvalho, da Universidade Federal do Rio de Janeiro, e colaboradores, na revista Zootaxa, de distribuição on line e gratuita, em 9 de maio de 2011.
O primeira icnoespécie de ovos fossilizados da América do Sul
Ovo fossilizado de crocodyliforme , foto de Carlos de Oliveira.
Ninhos de 70 milhões de anos foram encontrados nas proximidades da cidade de Jales, interior do estado de São Paulo, por Carlos de Oliveira, da Fundação Educacional de Fernandópolis (SP). A descoberta foi publicada este mês na revista Paleontoloy por ele e colaboradores. Os ovos foram encontrados em 2006 em rochas da Formação Adamantina, Grupo Bauru. A grande concentração deste material chamou a atenção de Carlos, que com o prosseguimento das escavações encontrou o que seria equivalente a 17 ninhadas situadoas em 3 diferentes níveis de deposição sedimentar (o que corresponderia a três eventos temporais diferentes).
Foto por Carlos de Oliveira.
Os ovos são alongados e tem forma elíptica. O tamanho varia entre 5,8 e 6,5 cm. A maioria estava quebrada, o que pode sugerir que os filhotes haviam nascido e deixado os ovos. Apenas alguns estavam completos.
Esqueleto parcial de Baurusuchus encontrado em associação com os ninhos. Foto: Carlos Oliveira.
Todos eles foram considerados como pertencentes a um gênero de crocodyliforme, Baurusuchus, devido a inúmeros ossos, e inclusive crânios e esqueletos parciais destes animais, que foram encontrados associados aos ninhos. Mas não foi só isso que ajudou os pesquisadores a definirem os produtores destes ovos. A microestrutura da casca também revela detalhes sobre quem os depositou:
Ovos de aves, dinossauros, crocodyliformes, quelônios, lagartos e cobras têm estruturas macro e microscópicas diferentes. Além do tamanho e formato ovo, aspectos histoestruturais da casca, como a organização do sistema de poros e a forma de deposição de cálcio (existem diferentes morfotipos estruturais: testudinóide, crocodilóide, dinossauróide, ornitóide e geckóide por exemplo) ajudam a identificar o produtor.
Tipos de ovos de acordo com sua microestrutura. A micro-estrutura pode ser avaliada por meio de cortes histológicos da casca, que são então observados com o auxílio da microscopia eletrônica de varredura.
O estudo dos ovos fossilizados se chama Paleo-oologia e está inserida numa área da paleontologia chamada de Paleoicnologia, palaios=antigo, iknos=vestígios e logos=estudo, ou seja, O estudo dos vestígios antigos.
A Paleoicnologia estuda todo tipo de vestígio fóssil indireto ou evidência comportamental de uma atividade biológica (produzida por um organismo extinto). Os ovos constituem vestígios do comportamento de reprodução de animais extintos, logo estão no escopo de estudo da Paleoicnologia, assim como as pegadas fósseis, por exemplo, que são vestígios de locomoção.
A Paleoicnoloia tem toda uma taxonomia própria para definir diferentes tipos de vestígios. Essa ‘parataxonomia’, à modo do sistema de nomenclatura biológica, é binomial e latinizada. Se as características gerais de uma estrutura paleoicnológica foram parecidas com as de materiais já conhecidos, elas recebem o mesmo nome destes, mas se foram diferentes, ganham uma nova designação, como uma nova espécie. A propósito, ICNOespécie e icnogênero são a maneira correta de se denominar estas estruturas, para não se confundir com o sistema de nomenclatura biológica – o que é muito comum.
Por exemplo, o icnogênero de pegadas de mamíferos conhecido como Brasilichnium elusivum ( descrito para Fm. Botucatu, Bacia do Paraná) comumente é confundido com o nome do produtor das pegadas… que na verdade não é conhecido por nenhuma evidência de fóssil corporal! O nome B. elusivum se refere somente às pegadas, não ao seu produtor. Até mesmo animais diferentes poderiam ter produzido o mesmo tipo de vestígio. Cuidado…
No caso do material de Jales, SP, os autores consideraram que todas as características identificadas poderiam sustentar um novo icnogênero, que denominaram de Bauruoolithus fragilis. Esta seria a primeira icnoespécie de ovos fossilizados descrita para a América do Sul (mas não os primeiros ovos fósseis descritos nem para o Brasil, nem para a América Latina! Há abundantes registros de ovos fossilizados na Argentina e vários também aqui no Brasil. Referências em nosso país são os ovos de dinossauro encontrados na região de Uberaba e os ovos atribuídos a Mariliasuchus em Marília, SP).
As feições encontradas nos ovos, segundo os autores, são muito diferentes daquelas encontradas em outros crocodilomorfos, o que leva a suspeita de que os produtores de Bauruoolithus teriam um modo de reprodução peculiar. Isto pode estar diretamente ligado com o sucesso ecológico do grupo e pode fornecer respostas interessantes quanto a adaptação destes animais às condições ambientais do sudeste brasileiro durante o Cretáceo Tardio.
O estudo de ovos fósseis pode revelar detalhes de aspectos biológicos e ecológicos dos seus produtores. –estratégias ou comportamentos de reprodução estão intimamente ligadas ao rigor ambiental e estresse ecológico (competição, predação, etc), assim como aspectos paleoambientais – recuperados direta (tafonomia) ou indiretamente (um estresse ambiental -uma grande seca, período de escassez de alimentos, etc. – pode ser detectado estudando-se a microestrutura dos ovos)-, e paleoclimáticos(inferidos utilizando-se análise de isótopos). Estes estudos são um passo além da simples descrição.
Depois do anúncio de Tapuiasaurus em fevereiro, somam-se à lista de vertebrados mesozóicos do Brasil o gigante dino carnívoro Oxalaia, mais um bizarro crocodilomorfo terrestre – Pepesuchus – e Brasiliguana, um pequeno lagarto da região de Presidente Prudente, SP…
Mas isso não é só, ainda há muito mais por vir!!
Tapuiasaurus foi descrito ainda em fevereiro na revista científica de divulgação livre, PLoS ONE (acesse o artigo aqui). O anúncio do bicho foi um sucesso: Dinossauro na cabeça! Um crânio completo foi apresentado por Zaher e colaboradores e o estado de conservação do material deixou pesquisadores do mundo inteiro boquiabertos. Tapuiasaurus pertence a um grupo de dinossauros chamado de saurópodes (dinos gigantes de pescoço e cauda longos) e mais especificamente a um ramo chamado de ‘titanossaurídeos’. Crânios de dinossauros saurópodes são relativamente raros, já que tendem a logo se desarticular do corpo depois da morte do animal. Por isso Tapuiasaurus foi recebido com tanta festa.!
A idade do fóssil está entre 125 e 112 milhões de anos. Ele foi encontrado nos estratos cretácicos da Bacia Sanfranciscana, nas imediações do município de Coração de Jesus, Minas Gerais, próximo a divisa com a Bahia.
Não só o crânio, mas vértebras, partes da escápula, um rádio e um fêmur também foram descritos.
O crânio é impressionante. Com o focinho alongado e a abertura nasal na altura dos olhos, ele lembra aquele de outros titanossauros como Rapetosaurus e Nemegtosaurus. Porém, Tapuiasaurus viveu bem antes destes animais – pelo menos 30 milhões de anos antes. Em termos evolutivos, essa é uma informação muito importante. Tudo indica que este formato craniano, comumente encontrado em dinossauros saurópodes titanossaurídeos do final do Período Cretáceo, já havia evoluído bem antes do que se pensava.
Uma exposição temática com os fósseis do animal está sendo apresentada no Museu de Zoologia da USP em São Paulo. Vale a pena visitar!! Exposição “Cabeça Dinossauro”.
Quanto a Oxalaia, anunciado à imprenssa brasileira no dia 16 de março, temos um registro bem menos impressionante, mas tão importante quanto o de Tapuiasaurus. Oxalaia tratava-se de um imenso dinossauro carnívoro espinossaurídeo (da família do Espinossauro e do Suchomimus, dinos com o focinho alongado e achatado como o dos crocodilos), que devia medir entre 12 e 14 metros. Seria o segundo maior dinossauro dessa família de terópodes. Os restos do animal foram encontrados ainda em 1999 durante uma expedição da equipe de paleontólogos do Museu Nacional à Ilha do Cajual, no Estado do Maranhão (Leia aqui!). Encontrado na famosa ‘Laje do Coringa’, o nível mais fossilífero da Formação Alcântara, o bicho parece ter sido um elemento comum no ambiente pretérito daquela região, onde são encontrados abundantes dentes do animal. Foram descritos por Kellner e colaboradores dois fragmentos de crânio, considerados suficientes para definir a nova espécie. O trabalho foi apresentado numa edição especial dos Anais da Academia Brasileira de Ciências e pode ser acessado aqui. Oxalaia pode ser considerado hoje o maior dinossauro carnívoro brasileiro. Três espécies de espinossaurídeos já foram descritas para o Brasil: Irritator challengeri, Angaturama limai e Oxalaia quilombensis. O nome Oxalaia faz referência a divindade africana Oxalá e quilombensis à um antigo Quilombo da região da Ilha do Cajual.
Fragmentos do crânio de Oxalaia, descritos por Kellner e colaboradores.
Reconstituição artística do animal por Maurílio de Oliveira.
Pepesuchus pode parecer um nome estranho, mas foi uma homenagem ao Prof. José Martin Suaréz (conhecido por seus colegas como Pepe) para nomear o mais novo crocodilomorfo terrestre barsileiro. Descrito por Diógenes de Almeida Campos e colaboradores, a nova espécie conta com dois crânios quase completos e mandíbulas. O material é proveniente do famoso sítio “Tartaruguito” (Fm. Presidente Prudente, Grupo Bauru, Bacia Bauru), próximo às cidades de Pirapozinho e Presidente Prudente, no Estado de São Paulo. A nova espécie foi classificada como um peirossaurídeo e acrescenta ainda mais ao conhecimento desses animais na Bacia Bauru, Cretáceo Superior brasileiro. Os peirossaurídeos parecem ter sido um dos clados de Mesoeucrocodylia mais bem representados no paleocontinente austral, Gondwana. O material pós-craniano da nova espécie será descrito separadamente.
Reconstituição do crânio de Pepesuchus.
Reconstituição artística de Pepesuchus.
Por fim, não poderíamos deixar de falar de Brasiliguana, também publicado na edição especial dos Anais da Academia Brasileira de Ciências (acesse aqui). Brasiliguana trata-se de um pequeno lagarto dos estratos do Cretáceo Superior da Formação Adamantina, Bacia Bauru, da região do município de Presidente Prudente, SP. O registro de squamatas no Brasil é raro e inclui somente 6 apontamentos: Tijubina,Olindalacerta e squamata indeterminado, da Bacia do Araripe, e Pristiguana e 2 registros também não específicos da Bacia Bauru. Brasiliguana viria a acrescentar este conhecimento.
O animal foi descrito por William Nava e Agustín Martinelli com base em um fragmento cranial, cujos formato e implantação dos dentes, de acordo com os autores, são semelhantes a dos lagartos iguanídeos.
Material descrito de Brasiliguana prudentensis.
Já que falamos tanto da edição especial dos Anais da Academia Brasileira de Ciências, vale a pena checar os outros artigos. Você os encontra disponíveis aqui.
Não deixe de dar uma olhada naquele de Bittencourt & Langer (aqui). O amigo Johnny fez uma fantástica revisão sobre as ocorrências de dinossauros no Brasil e as suas relações biogeográficas. Referência!
As novidades por enquanto são estas, mas fiquem de olho porque tem muito mais por vir!



 O fóssil de Gondwanasuchus scabrosus é representado por um crânio parcialmente completo, que foi encontrado em 2008 em associação a um grande indivíduo de Baurusuchus salgadoensis, um crocodiliforme também da família dos baurussuquídeos. Gondwanasuchus scabrosus convivia não só com outros baurussuquídeos, mas também com crocodiliformes herbívoros da família dos esfagessaurídeos. A presença de esfagessaurídeos e o fato de os depósitos da Formação Adamantina no município de General Salgado serem basicamente compostos por paleossolos (solos que foram preservados no registro geológico), sugerem que pelo menos algumas partes do habitat de Gondwanasuchus eram compostas por áreas com vegetação arbustiva e arbórea.
O fóssil de Gondwanasuchus scabrosus é representado por um crânio parcialmente completo, que foi encontrado em 2008 em associação a um grande indivíduo de Baurusuchus salgadoensis, um crocodiliforme também da família dos baurussuquídeos. Gondwanasuchus scabrosus convivia não só com outros baurussuquídeos, mas também com crocodiliformes herbívoros da família dos esfagessaurídeos. A presença de esfagessaurídeos e o fato de os depósitos da Formação Adamantina no município de General Salgado serem basicamente compostos por paleossolos (solos que foram preservados no registro geológico), sugerem que pelo menos algumas partes do habitat de Gondwanasuchus eram compostas por áreas com vegetação arbustiva e arbórea. Os dentes posteriores de G. scabrosus são altamente comprimidos e com bordas serrilhadas, como os dentes de alguns dinossauros carnívoros. Outra peculiaridade da dentição desses animais é a presença de profundas estrias que percorrem os dentes da base para o topo, possivelmente garantindo uma maior resistência a quebra durante o processo de caça e alimentação.
Os dentes posteriores de G. scabrosus são altamente comprimidos e com bordas serrilhadas, como os dentes de alguns dinossauros carnívoros. Outra peculiaridade da dentição desses animais é a presença de profundas estrias que percorrem os dentes da base para o topo, possivelmente garantindo uma maior resistência a quebra durante o processo de caça e alimentação.

















{kind=link}