

Esta contribuição foi feita pelo aluno de graduação da Universidade Federal de Uberlândia (UFU) chamado Rodolfo Otávio dos Santos. Atualmente ele se encontra no 6º período do curso de Ciências Biológicas e está estagiando no Laboratório de Paleontologia da UFU (https://www.facebook.com/PaleoUFU).
Devido ao seu interesse na área e de auxiliar na divulgação sobre tais assuntos, ele seguirá contribuindo com mais postagens sobre os mais variados tópicos. Rodolfo fez sua primeira contribuição tratando sobre a discrepância entre a abundância de dinossauros na Argentina x Brasil (aqui) e sua segunda discutindo sobre quais fósseis reais foram usados como base para os Pokemon fósseis (aqui). Hoje elecomenta brevemente sobre uma ciência que não é nova, mas que nas últimas décadas tem apresentado novas descobertas incríveis. A ciência em questão é a Astrobiologia, bastante em voga nas ultimas semanas devido a descoberta de novos planetas com potencial significativo de haver vida!
Uma forma bastante eficiente de se conhecer uma ciência é saber qual seu objeto de estudo, ou seja, quais perguntas ela busca responder. Dessa forma a Biologia, por exemplo, tem como objetivo final explicar a totalidade das questões relacionadas à vida. Para áreas emergentes da ciência, no entanto, delimitar as questões que as concernem não é uma tarefa muito simples, pois a maioria aborda temas cujo nosso conhecimento atual é demasiadamente pequeno. Podemos citar, como exemplo, estudos sobre a astropaleobiologia, o tema deste texto.
É preciso, de início, definir o termo astropaleobiologia, que é: área da ciência responsável pelo estudo dos fósseis encontrados fora do planeta Terra. Nesse sentido, é válido mencionar que o vocábulo astropaleontologia já era utilizado anteriormente, porém com outros significados, como: o estudo da evolução das estrelas e/ou o estudo de como os eventos astronômicos influenciaram a vida da Terra. Sendo assim, o termo astropaleobiologia é atualmente o mais utilizado para designar a área que estuda os restos de organismos vivos que porventura habitaram outros locais do nosso universo.
A partir do momento que os astrônomos começaram a estimar com maior precisão o tamanho do universo e se depararam com sua grandeza, logo perceberam que as escalas de medidas convencionais eram ineficazes para as distâncias cósmicas. A noção de que o universo é infinitamente grande e antigo fez com que novos questionamentos surgissem. Parece existir uma incongruência entre a quantidade de espaço disponível e o número de formas de vida no universo, conhecida popularmente como Paradoxo de Fermi. Em outras palavras, parafraseando o célebre astrônomo norte americano Carl Sagan: “Seria o universo um grande desperdício de espaço?”.

Quando o assunto é a possibilidade de vida extraterrestre, outro tópico importante é a Equação de Drake. Trata-se de um famoso cálculo, criado por Frank Drake, que busca estimar a quantidade de civilizações presentes na galáxia, partindo do uso de algumas variáveis (até então impossíveis de serem mensuradas na época de sua criação, ainda que hoje algumas delas sejam razoavelmente conhecidas). A equação foi recentemente atualizada, havendo uma substituição de algumas variáveis por outras que atualmente são capazes de serem mensuradas, passando a ser conhecida como Equação de Seager.

A descoberta de organismos vivos fora da Terra traria implicações para toda nossa sociedade, principalmente para a ciência, filosofia e religião. Por exemplo, a possibilidade de múltiplas origens do que conhecemos como vida, ou o fato dos organismos vivos terem se originado em outros locais do universo e posteriormente terem colonizado a Terra (a famosa panspermia cósmica) revolucionariam toda a Sistemática Filogenética e, consequentemente, o modo como entendemos as relações de parentesco entre os seres vivos.
Mesmo nos dias atuais, não existe um consenso entre os biólogos sobre uma definição universal de vida. A existência de seres extraterrestres, que provavelmente teriam uma bioquímica muito diferente da nossa, poderia fazer com que a resposta para tal pergunta ficasse ainda mais difícil, pois ampliaria o leque de possibilidades para aquilo que definimos como um ser vivo. Em Titã, um dos satélites naturais de Saturno, cientistas têm especulado sobre um possível tipo de vida muito diferente da terrestre, baseado em hidrocarbonetos como o metano, dada a ausência de água líquida nessa lua.

Do ponto de vista astropaleobiológico, a hipótese mais interessante seria a de que, tal como ocorre na Terra, as taxas de extinções de seres vivos no universo sejam grandes, de forma que a grande maioria dos organismos já se extinguiram. Dessa forma, para conhecermos de fato essa diversidade inimaginável, teríamos que estudar os vestígios por eles deixados, provavelmente análogos ao que conhecemos como “fósseis”. Portanto, nesse momento, entra em cena a Astropaleobiologia.
Desde o final do século XX, graças às melhorias em nossa tecnologia, foi possível detectar os primeiros exoplanetas (planetas localizados fora do sistema solar). Atualmente, são conhecidos mais de 3000, alguns deles com características semelhantes às encontradas na Terra e consequentemente, os locais mais prováveis de encontrarmos vida fora de nosso planeta. Os radiotelescópios também têm ajudado os astrobiólogos na procura pela vida extraterrestre. Em 2016, a China inaugurou o maior até então já construído, o que pode ser um passo definitivo para respondermos a questão: estamos ou não sozinhos no universo?

Apesar do pequeno número de evidências, existem materiais para estudos astropaleobiológicos e, por mais paradoxal que possa parecer, esses materiais foram encontrados em nosso próprio planeta, porém suas origens remontam a um local distante. No passado, corpos celestes chocaram-se contra Marte, fazendo com que rochas marcianas fossem lançadas para o espaço. Eventualmente, algumas delas caíram na Terra e os cientistas, ao estudarem sua composição, perceberam que tais rochas não eram terrestres.
Em 1996, a notícia de que um meteorito (ALH 84001), encontrado na Antártica, continha estruturas muito semelhantes a fósseis de “bactérias marcianas” correu o mundo. Em 2006 cientistas analisaram outro meteorito, encontrado em 1911 no Egito, que possuía estruturas microtubulares, possível evidência de atividade microbiana. Mais recentemente, em 2014, outra rocha marciana (Y000593), encontrada no Japão, ganhou as manchetes por também apresentar microtúbulos, além de pequenas esferas, prováveis resquícios da presença de organismos vivos.
Alguns cientistas alegam que os microtúbulos seriam, na realidade, túneis escavados por estes organismos extraterrestres enquanto se alimentavam, de forma muito semelhante ao que é feito por algumas bactérias terrestres. Outros pesquisadores, entretanto, afirmam que tais estruturas teriam uma origem totalmente abiótica, sendo resultantes de reações físico-químicas desvinculadas de atividade biológica, pois não foram encontrados vestígios de moléculas capazes de se replicar. Há ainda a possibilidade de contaminação do material por organismos terrestres, o que dificulta os estudos.


Porém, de nada adianta os organismos vivos deixarem restos de sua existência para trás se os futuros cientistas não conseguirem ter acesso aos materiais. Em nosso planeta, conseguimos ter acesso aos fósseis pois as camadas em que eles se encontram são soerguidas graças à forças vindas do interior da Terra, possibilitando aos paleontólogos acesso mais fácil aos materiais. Isso só é possível devido ao fato de que a Terra é um planeta geologicamente ativo. No sistema solar, corpos celestes como Vênus provavelmente compartilham essa característica, enquanto outros, como Mercúrio e Marte, são geologicamente inativos, fator que dificultaria, e muito, o trabalho dos futuros astropaleobiólogos.
É importante salientar que as buscas por vida extraterrestre são altamente enviesadas, pois nossa procura se concentra em locais semelhantes à Terra (restrita, portanto, a planetas rochosos e com água). De forma similar, a procura pelos “fósseis” extraterrestres também está limitada ao nosso conhecimento acerca dos processos de fossilização terrestres. Entretanto, muito provavelmente, a imensa biodiversidade universal, ainda oculta, deve carregar consigo uma gama ainda maior de processos que desafiam nosso conhecimento.

A busca por organismos extraterrestres, estejam eles já extintos ou ainda vivos, é sobretudo uma forma de conhecermos a nós mesmos, de entendermos qual nosso papel no universo. No futuro, talvez o conhecimento adquirido com o estudo de possíveis “fósseis” extraterrestres, possamos definir e explicar de forma mais satisfatória o fenômeno que denominamos de vida. Quanto aos paleontólogos do presente, resta usar a imaginação, na tentativa de vislumbrar o passado de outros mundos, e aguardar pacientemente o progresso da ciência em sua procura por organismos extraterrestres.
Adendo: Duas descobertas recentes trouxeram grandes avanços para a Astropaleobiologia. A primeira pesquisa revelou a existência de um sistema composto por sete planetas rochosos, distante 39 anos-luz da Terra, orbitando a estrela TRAPPIST-1, dos quais três estão na chamada zona habitável, a região em que, caso exista água, ela se encontra no estado líquido, aumentando as chances de encontrarmos organismos vivos. Foi a primeira vez que um sistema contendo tantos planetas com grande potencial para abrigar vida foi encontrado.
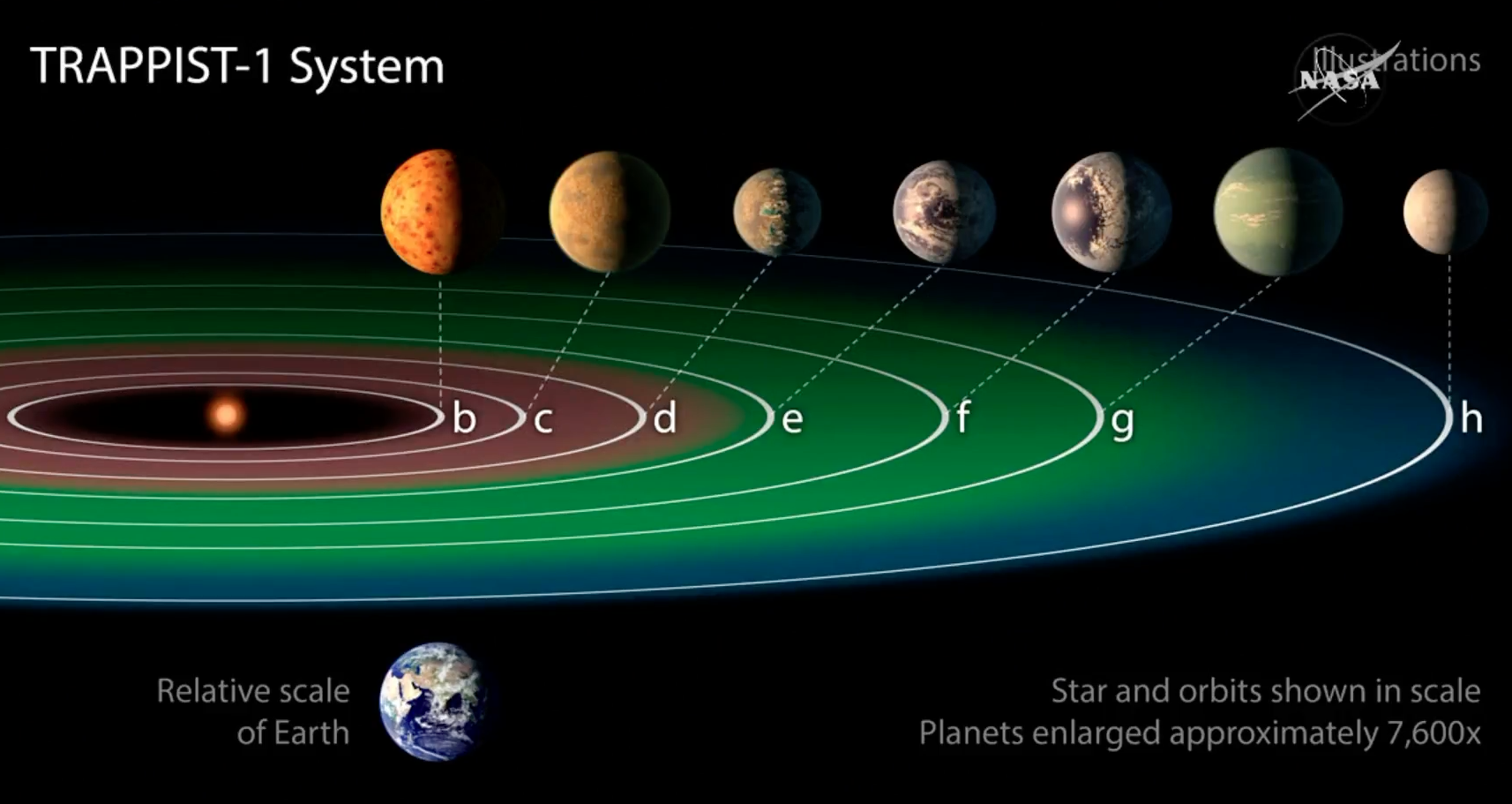
A segunda pesquisa mostrou a existência de fósseis de bactérias com uma idade entre 3,8 e 4,3 bilhões de anos, os mais antigos encontrados até o momento. Os materiais foram encontrados no Nuvvuagittuq Supracrustal Belt, em Quebec. No passado, este local foi um sistema de fontes hidrotermais rico em ferro, elemento que era utilizado no metabolismo dessas bactérias, que deixaram vestígios na forma de pequenos túbulos. Essa descoberta indica que a vida na Terra apareceu pouco tempo após a formação dos oceanos.

Os autores do estudo ainda lembraram que as condições do planeta Marte há 4,3 bilhões de anos eram semelhantes às da Terra primitiva, um forte indício de que a vida possa ter prosperado também no planeta vermelho, ainda que por um curto período de tempo. Considerando tais estudos, a existência de vida extraterrestre ganhou fortes evidências a seu favor e agora é uma questão e tempo até que novas descobertas sobre o assunto sejam encontradas, confirmando a existência de vida extraterrestre.
Referências Bibliográficas:
Astropaleobiologia:
COX, G. Astropaleobiology. Disponível em: <https://starscapescientific.wordpress.com/2012/06/09/astropaleobiology/>. Acesso em 4 de mar. 2017.
Paradoxo de Fermi:
NUNES, J. O Paradoxo de Fermi. Disponível em: <http://www.universoracionalista.org/o-paradoxo-de-fermi/>. Acesso em 4 de mar. 2017.
Equação de Drake e Seager:
OLIVEIRA, D. R. A. Equação de Drake para a vida alienígena recebe um upgrade. Disponível em: <http://www.universoracionalista.org/equacao-de-drake-para-a-vida-alienigena-recebe-um-upgrade/>. Acesso em 4 de mar. 2017.
PONTES, F. A caçadora de extraterrestres: A exótica missão da astrônoma Sara Seager, em busca de planetas habitáveis pelo Universo. Disponível em: <http://epoca.globo.com/vida/noticia/2013/08/cacadora-de-bextraterrestresb.html>. Acesso em 4 mar. 2017.
Vida em Titã:
HRALA, J. Life “Not as We Know It” Might Be Possible on Titan. Disponível em: <http://www.sciencealert.com/life-on-titan-might-be-completely-different-than-the-life-we-re-familiar-with>. Acesso em 4 de mar 2017.
Radiotelescópio Chinês:
O’NEILL, I. Monster Chinese Telescope the Next ET Hunter?. Disponível em: <http://www.seeker.com/monster-chinese-telescope-the-next-et-hunter-1765285433.html>. Acesso em 4 de mar. 2017.
Vida em Marte e Meteoritos:
SÉRVULO, F. Como procurar por vida em Marte?. Disponível em: <http://www.universoracionalista.org/como-procurar-por-vida-em-marte>. Acesso em: 4 de mar. 2017.
MCKAY, D. S. et al. Search for past life on Mars: Possible relic biogenic activity in martian meteorite ALH84001. Science, Washington, v. 273, p. 924-930, ago. 1996.
MIKOUCHI, T. et al. Mineralogy and petrology of Yamato 000593: Comparison with other Martian nakhlite meteorites. Antarctic Meteorite Research, Washington, v. 16, p. 34-57, fev. 2003.
MCKAY, D. S. et al. Life on Mars: new evidence from martian meteorites. Proceedings of SPIE Annual Meeting, Bellingham, v. 7441, p. 80-102, ago. 2009.
WARMFLASH, D.; WEISS, B. Dis life come from another world?. Disponível em: <http://www.bibliotecapleyades.net/ciencia/esp_ciencia_life09.htm>. Acesso em 4 de mar. 2017.
Missão Kepler:
NASA. Importance of Planet Detection. Disponível em: <https://kepler.nasa.gov/Mission/QuickGuide/>. Acesso em 4 de mar. 2017.
CLARK, S. Kepler space telescope in emergency mode. Disponível em: <https://spaceflightnow.com/2016/04/09/kepler-space-telescope-in-emergency-mode/>. Acesso em 4 de mar. 2017.
Exoplanetas:
SCHNEIDER, J. et al. Defining and cataloging exoplanets: the exoplanet.eu database. Astronomy & Astrophysics. Paris, v. 532, p. 79-90, jul. 2011.
LOPES, M. Métodos de Detecção de Planetas Extrasolares. Disponível em: <http://www.astropt.org/2013/11/20/metodos-de-deteccao-de-planetas-extrasolares/>. Acesso em 4 de mar. 2017.
Planetas geologicamente ativos:
SANCEVERO, S. Vênus é um planeta geologicamente ativo?. Disponível em: <http://www.astropt.org/2016/11/30/venus-e-um-planeta-geologicamente-ativo/>. Acesso em: 4 de mar. 2017.
Planetas em TRAPPIST-1:
BARSTOW, J. K.; IRWIN, P. G .J. Habitable worlds with JWST: transit spectroscopy of the TRAPPIST-1 system?. Monthly Notices of the Royal Astronomy Society, Oxford, v. 461, n. 1, p. 92-96, mai. 2016.
Fósseis mais antigos já encontrados:
DODD, M. S. et al. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature, Londres, v. 543, p. 60-64, jan. 2017.













 Tiago Rodrigues Simões possui graduação e mestrado em Ciências Biológicas pela Universidade Federal do Rio de Janeiro e atualmente está concluindo o doutorado na University of Alberta (Edmonton, Canandá). A sua pesquisa consiste no estudo da origem e evolução de Squamata (lagartos e cobras), utilizando dados de espécies fósseis e viventes (
Tiago Rodrigues Simões possui graduação e mestrado em Ciências Biológicas pela Universidade Federal do Rio de Janeiro e atualmente está concluindo o doutorado na University of Alberta (Edmonton, Canandá). A sua pesquisa consiste no estudo da origem e evolução de Squamata (lagartos e cobras), utilizando dados de espécies fósseis e viventes (







 Nomes: Omanyte e Omastar (estes nomes são uma derivação de Amonite, sendo que em Omastar há também uma referência ao formato de estrela star, devido a forma dos tentáculos do Pokémon).
Nomes: Omanyte e Omastar (estes nomes são uma derivação de Amonite, sendo que em Omastar há também uma referência ao formato de estrela star, devido a forma dos tentáculos do Pokémon).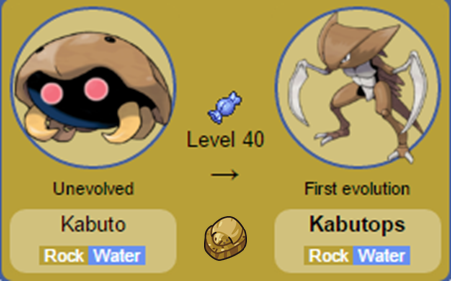 Nomes: Kabuto e Kabutops (Kabuto é o nome dado aos capacetes utilizados pelos samurais).
Nomes: Kabuto e Kabutops (Kabuto é o nome dado aos capacetes utilizados pelos samurais).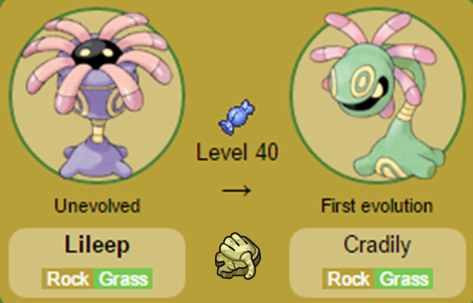 Nomes: Lillep e Cradily (Do inglês Lily, pois foram baseados nos Lírios do mar).
Nomes: Lillep e Cradily (Do inglês Lily, pois foram baseados nos Lírios do mar). Nomes: Anorith e Armaldo (Anorith é a junção de Anomalocaris, animal no qual o Pokémon foi baseado e lith, sufixo que significa rocha; enquanto Armaldo é uma referência à Armor, armadura em inglês).
Nomes: Anorith e Armaldo (Anorith é a junção de Anomalocaris, animal no qual o Pokémon foi baseado e lith, sufixo que significa rocha; enquanto Armaldo é uma referência à Armor, armadura em inglês).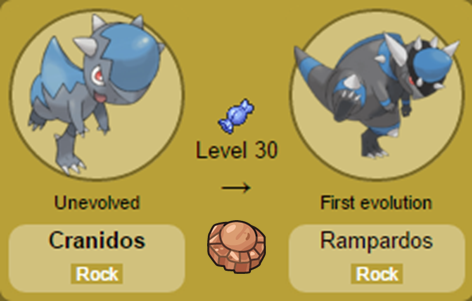 Nomes: Cranidos e Rampardos (Cranidos é derivado de Crânio, característica marcante dos Pachycefalosaurus, os dinossauros que serviram de inspiração aos Pokémon, enquanto Rampardos pode ser uma referência à palavra Rampage, que significa “ficar com raiva” em português).
Nomes: Cranidos e Rampardos (Cranidos é derivado de Crânio, característica marcante dos Pachycefalosaurus, os dinossauros que serviram de inspiração aos Pokémon, enquanto Rampardos pode ser uma referência à palavra Rampage, que significa “ficar com raiva” em português). Nomes: Shieldon e Bastiodon (O nome Shieldon é derivado de shield, escudo em inglês, enquanto Bastiodon vem da palavra bastion, baluarte em português, uma construção defensiva cercada por muralhas, muito utilizada na Europa a partir do séc. XV).
Nomes: Shieldon e Bastiodon (O nome Shieldon é derivado de shield, escudo em inglês, enquanto Bastiodon vem da palavra bastion, baluarte em português, uma construção defensiva cercada por muralhas, muito utilizada na Europa a partir do séc. XV).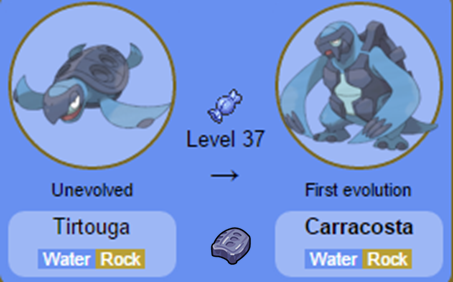 Nomes: Tirtouga e Carracosta (O nome Tirtouga é baseado em Tortuga, tartaruga em espanhol; enquanto Carracosta provavelmente faz referência à carapaça característica do grupo e à costa litorânea, local de nascimento das tartarugas marinhas).
Nomes: Tirtouga e Carracosta (O nome Tirtouga é baseado em Tortuga, tartaruga em espanhol; enquanto Carracosta provavelmente faz referência à carapaça característica do grupo e à costa litorânea, local de nascimento das tartarugas marinhas).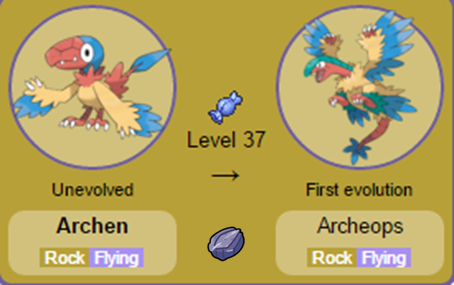 Nomes: Archen e Archeops (O nome dos dois Pokémon é baseado em Archaeopteryx, um dinossauro muito semelhante às aves, que possuía asas, bico e penas).
Nomes: Archen e Archeops (O nome dos dois Pokémon é baseado em Archaeopteryx, um dinossauro muito semelhante às aves, que possuía asas, bico e penas). Nomes: Tyrunt e Tyrantrum (Tyrunt é a combinação entre Tyrannosaurus e runt, palavra em inglês utilizada para designar alguém pequeno; enquanto Tyrantrum é a combinação de Tyrannosaurus e tantrum, palavra também de origem inglesa que significa ataque de raiva).
Nomes: Tyrunt e Tyrantrum (Tyrunt é a combinação entre Tyrannosaurus e runt, palavra em inglês utilizada para designar alguém pequeno; enquanto Tyrantrum é a combinação de Tyrannosaurus e tantrum, palavra também de origem inglesa que significa ataque de raiva). Nomes: Amaura e Aurorus (O nome de ambos é baseado na junção de duas palavras: Amargasaurus, o dinossauro em que foram baseados, e aurora, uma referência ao fenômeno que ocorre na atmosfera, em regiões próximas aos polos).
Nomes: Amaura e Aurorus (O nome de ambos é baseado na junção de duas palavras: Amargasaurus, o dinossauro em que foram baseados, e aurora, uma referência ao fenômeno que ocorre na atmosfera, em regiões próximas aos polos).