
Olá caros leitores, depois de um breve período de pausa nas postagens, voltamos com grande estilo. Hoje apresento a vocês um interessante texto redigido pelo Mestrando em Zoologia do Museu Nacional/UFRJ Geovane Alves de Souza, O assunto abordado se refere as primeiras discussões históricas sobre as hipóteses filogenéticas dos Amniotas (grupo que tradicionalmente inclui os répteis, aves e mamíferos e suas formas relacionadas) e a importância dos fósseis para um melhor entendimento da evolução das espécies. Então, sem mais delongas, vamos ao texto!
Editado em 19/08/2018.
Em plena Era da Filogenômica, na qual o DNA possui papel central na busca pela compreensão da evolução da vida na Terra, é comum pensarmos nos fósseis como uma fonte de dados um tanto quanto ultrapassada, trabalhosa e que demanda muito tempo para estudar. A diretora do Jurassic World, Claire Dearing (encenada pela atriz Bryce Dallas) enfatiza isso muito bem em sua fala: “Aprendemos mais com a genética em 10 anos do que em um século, escavando”. Hoje é consenso no meio acadêmico a importância dos vestígios da vida pretérita, os fósseis (para melhores detalhes acerca de sua definição veja aqui), quando inferimos relações de parentesco dos organismos viventes. Contudo, nem sempre os fósseis tiveram sua importância reconhecida e passando por momentos de glória e queda ao longo dos últimos séculos.
Após a publicação da obra A Origem das Espécies por Meio da Seleção Natural por Charles Darwin em 1859, no qual o autor defendia as teorias de Evolução Biológica, Seleção Natural e Ancestralidade Comum, os fósseis passaram a desempenhar um papel chave na compreensão de como a vida evoluiu. Contudo, foi na metade do século XX, que o registro fossilífero enfrentou uma queda brusca de sua supremacia. O responsável foi o advento de uma nova maneira de se estudar a evolução: a Sistemática Filogenética de Willi Hennig (1950). A nova metodologia e filosofia da sistemática nos seus primeiros anos de existência não exigia a necessidade de determinar uma dada espécie fóssil conhecida como ancestral entre duas linhagens. Podíamos estudar a evolução das espécies viventes, tratando o ancestral comum entre elas como uma espécie hipotética. Esse modo de vislumbrar as árvores filogenéticas (diagramas ramificados que representam a evolução de uma linhagem e que são gerados a partir de uma análise computacional ou análise filogenética), conhecido como Modelo Cladogenético, é antagônico ao antigo modelo vigente, o Anagenético. De fato, determinar em qual ponto exato da evolução de uma linhagem uma espécie fóssil esta inserida não é uma tarefa fácil, eu diria que um tanto quanto impossível, a menos que tenhamos uma máquina do tempo para voltarmos e acompanharmos o passo a passo da evolução de determinada linhagem ao longo dos milhares de anos. Muitas críticas contra o uso dos fósseis foram levantadas na época. O próprio Hennig reconhecia que os dados fósseis poderiam ser úteis na hora de conduzir uma análise filogenética. Contudo, devido à tamanha incompletude do registro fossilífero, ou seja, tanta informação biológica era perdida no processo de formação de um fóssil, que estes deveriam ser preferivelmente menos utilizados na hora de reconstruir as relações de parentesco.
Petterson (1981) mostrou, através de vários exemplos, o quanto os fósseis prejudicavam a compreensão sobre as hipóteses de evolução dos animais. Ax em 1987 defendeu em seu livro The Phylogenetic System que os dados fósseis são tão incompletos que as árvores deveriam ser construídas com base apenas nos grupos viventes e só depois que a análise computacional fosse feita é que se deveriam adicionar os fósseis. Dessa maneira e com muito sucesso, os críticos rapidamente conseguiram marginalizar o uso dos dados paleontológicos nos estudos da evolução das linhagens de organismos viventes.
A Hipótese Clássica da evolução dos Amniotas
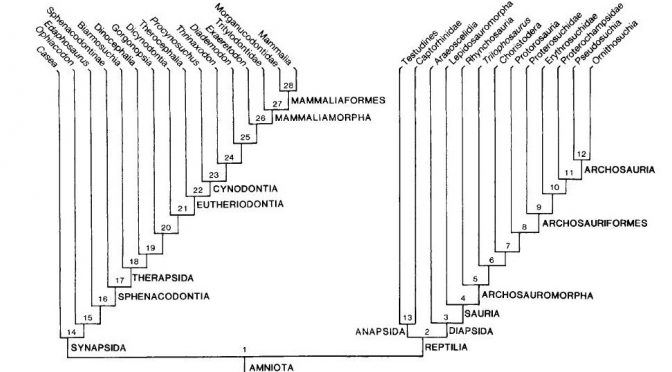
Inúmeras árvores foram construídas ao longo dos anos seguintes, a maioria delas ignorando as informações provindas dos fósseis. O estudo que mais me chamou a atenção foi o trabalho clássico de Gardiner em 1982, no qual este autor tentou reconstruir a até então, pouco compreendida história evolutiva dos amniotas. Amniota é um grupo de animais vertebrados que possuem, dentre muitas características, uma membrana extraembrionária ao redor do feto chamada de amnion, membrana a qual é fundamental para a independência da água do ambiente durante o desenvolvimento do filhote no ovo, permitindo que estes animais colonizassem completamente o habitat terrestre. Estamos falando então da maioria esmagadora de vertebrados terrestres (e os que secundariamente retornaram ao ambiente aquático) que dominaram a Terra: desde as formas extintas famosas como dinossauros, pterossauros, ictiossauros, plesiossauros até as espécies contemporâneas de tartarugas, crocodilos, lagartos, serpentes, aves e mamíferos.
Gardiner utilizou vários dados morfológicos dos cinco grupos de amniotas viventes em suas análises (tartarugas, lagartos, jacarés, mamíferos e aves). Gardiner observou que as aves e os mamíferos eram evolutivamente relacionados, sendo agrupados por uma série de características que eram adaptações às suas altas taxas metabólicas (metabolismo alto leva a uma temperatura corpórea alta, sendo estes animais equivocadamente chamados de animais de “sangue quente”).
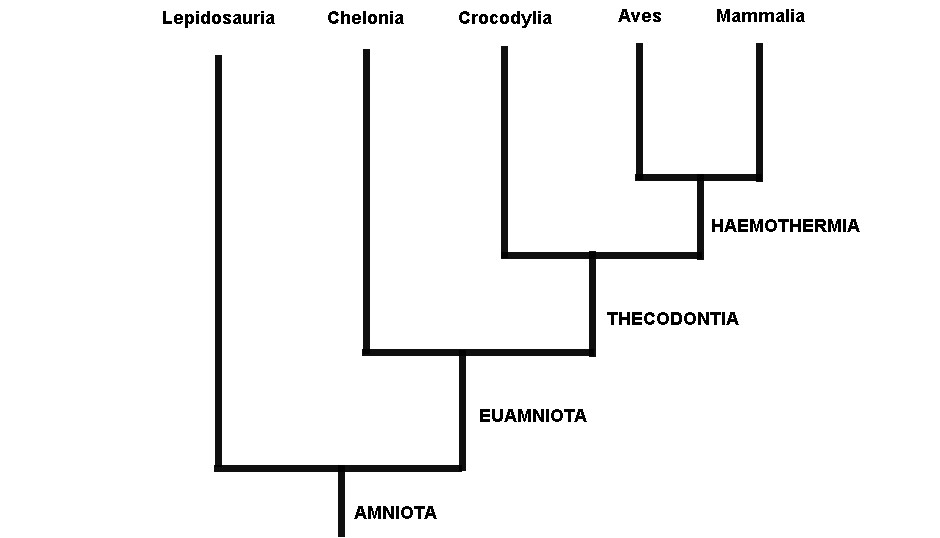
Gardiner ressuscitou o antigo termo Haeomothermia para nomear o grupo de animais de “sangue quente” formado por aves e mamíferos (o termo vem de homeotermia, do grego homo: igual, thermia: temperatura; que é como chamamos os animais que possuem temperaturas corporais constantes). Haemothermia, por sua vez era relacionado evolutivamente com o Crocodylia (crocodilos, jacarés e gaviais) formando o grupo Thecodontia. Tartarugas, cágados e jabutis (Chelonia) eram mais aparentados com Thecodontia (Crocodylia+(Aves+Mammalia)). Por fim, o grupo mais basal de Amniota era Lepidosauria, o qual abrange tuataras, serpentes e lagartos (Figura 1).
O trabalho experimental de Gauthier
Os resultados de Gardiner se baseam exclusivamente em animais viventes, conforme já foi dito e ecoaram por quase uma década. Lovtrup (1985) publicou um trabalho no qual afirmou ter encontrado maior suporte à hipótese de Gardiner. Até que, em 1988, um célebre manuscrito chegou para revolucionar a visão que a comunidade científica da época tinha sobre os dados paleontológicos. Gauthier e seus colegas (1988), baseados na hipótese de filogenia dos amniotas proposto por Gardiner, publicaram o primeiro estudo demonstrando empiricamente (ou seja, através de experimentos práticos e não calcados apenas em conjecturas) que os fósseis poderiam contribuir e muito na elucidação das hipóteses de parentesco dos seres vivos atuais. Para isso, eles conduziram uma nova análise, só que dessa vez incluindo espécies extintas, obtendo uma árvore marcadamente diferente e depois a submeteram a alguns testes para confirmar sua validade. Para entendermos como os autores chegaram a suas conclusões precisamos compreender os experimentos que a equipe realizou. Gauthier partiu de uma pergunta: os Fósseis poderiam alterar as nossas hipóteses de parentesco entre as biotas recentes? Para responder isso, ele conduziu um estudo em três etapas.
(1) Primeiramente, antes de rodar uma nova análise incluindo os fósseis, os pesquisadores destrincharam e reviram todas as características utilizadas por Gardiner e perceberam que havia alguns equívocos e erros na interpretação dos caracteres. Gardiner afirmara, por exemplo, que um coração dividido em quatro câmaras é homólogo (mesma origem; para melhor entendimento desta terminologia veja aqui) em crocodilos, aves e mamíferos, o que reforçava a relação de parentesco entre eles. Contudo, quando acompanhamos o desenvolvimento embrionário do septo interventricular destes animais, vemos que nos mamíferos ele se desenvolve a partir de uma crista de tecido endocárdico na parede de trás do ventrículo, enquanto que nas aves e nos crocodilos o septo surge de varias protuberâncias musculares pouco recobertas por endocárdio na lateral do ventrículo. Apesar de serem estruturas semelhantes nos adultos, elas não possuem a mesma origem no embrião, ou seja, não são homólogas. Logo, os equívocos nas interpretações de Gardiner o levaram a estabelecer homologias entre mamíferos e aves que não condiziam com a realidade.
(2) Depois de corrigir a lista de características, Gauthier rodou duas análises, uma contendo apenas os dados morfológicos dos cinco grupos viventes de Gardiner e outra incorporando 29 espécies de amniotas extintos. Foi então que uma nova hipótese começou a tomar forma.
A análise com dados dos animais viventes
A árvore filogenética obtida utilizando apenas os cinco grupos de amniotas viventes se assemelhou à hipótese clássica de Gardiner de 1982, com apenas uma pequena diferença: os crocodilos e não os mamíferos eram mais relacionados com as aves, trazendo dúvidas quanto a validade do antigo grupo Haemothermia (Ver figura 2). Isso implica em inferências importantíssimas para nossa compreensão da evolução da homeotermia. Primeiro, que um jacaré é o parente mais próximo das aves do que qualquer outro animal vivo hoje. Além disso, muitas características que antes eram vistas como homólogas entre aves e mamíferos e que estariam presentes no suposto ancestral comum destas duas linhagens, na verdade surgiram duas vezes independentemente na árvore da vida dos vertebrados. Talvez em resposta provavelmente as mesmas pressões evolutivas.
A análise com dados combinados (animais viventes + extintos)
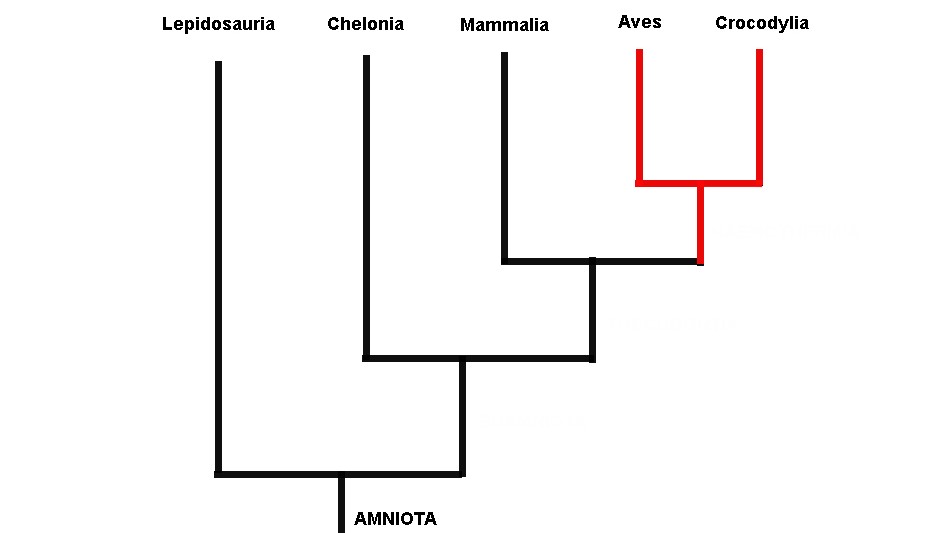
Na segunda análise na qual Gauthier acrescentou os fósseis, uma árvore completamente diferente surgiu (ver figura 3). Não só aves e crocodilos estavam agrupados juntos, mas lagartos e serpentes (Lepidosauria) agora estavam mais relacionados a Aves + Crocodylia do que as tartarugas. O mais estranho foi que os mamíferos agora estavam na base da árvore, formando uma grande dicotomia inicial: Mamíferos e todos seus parentes extintos de um lado versus lepidossauros, quêlonios, crocodilos, aves e todos seus parentes extintos relacionados do outro. Ao primeiro grupo, chamamos de Synapsida (amniotas que apresentam uma fenestra temporal no crânio) e ao segundo, Reptillia (que compreende tanto amniotas que possuem duas ou nenhuma fenestra temporal, Diapisida e Anapsida, respectivamente). Contudo, não bastava obter uma árvore completamente diferente, ela precisava ser mais bem justificada.
Análises posteriores – Esmiuçando a nova hipótese obtida
Na última etapa do estudo (3), uma série de experimentos computacionais foram conduzidos por Gauthier, alguns serão explicados mais adiante e que culminaram em um achado esperançoso, principalmente para nós paleontólogos: Fósseis são fundamentais para elaboração das hipóteses de relação entre as espécies viventes.
Gauthier e seus colegas queriam saber o porquê que as árvores diferiram tanto nas análises com e sem os fósseis. Para isso, eles fizeram algumas análises posteriores, contudo só irei detalhar três delas neste texto que acredito serem mais interessantes para nossa discussão. Primeiro, os autores compararam o índice de consistência de sua nova árvore com o índice da árvore de Gardiner. Este índice mostra o quão robusto e conciso estão seus resultados, no caso, sua hipótese de evolução de uma linhagem. O interessante foi que mesmo adicionando várias espécies fósseis na sua análise, Gauthier encontrou um valor de índice de consistência semelhante ao de Gardiner, mostrando que mesmo os fósseis alterando drasticamente a hipótese, a árvore continuava tão confiável quanto uma utilizando apenas espécies viventes.
Numa análise posterior, Gauthier e colaboradores removeram todos os grupos fósseis do lado “sinapsídeo” da árvore, deixando apenas os animais viventes desse ramo (mamíferos) junto com todos do lado “Reptillia” (viventes e extintos) e rodaram a análise. Paralelamente, foi feito o oposto, todas as linhagens extintas de Reptillia foram retiradas deixando apenas seus representantes viventes e o lado “sinapsídeo” da árvore (viventes e extintos) e rodaram a análise. Os autores viram que não importava qual fóssil de Reptillia fosse retirado, a árvore final não se alterava. Porém, quando os fósseis de sinapsídeos eram retirados, a árvore adquiria o novo padrão proposto por Gardiner (1982; Figura 3).
Gauthier então percebeu que os principais responsáveis pela nova topologia da árvore eram os fósseis de sinapsídeos. Mas por que isso?
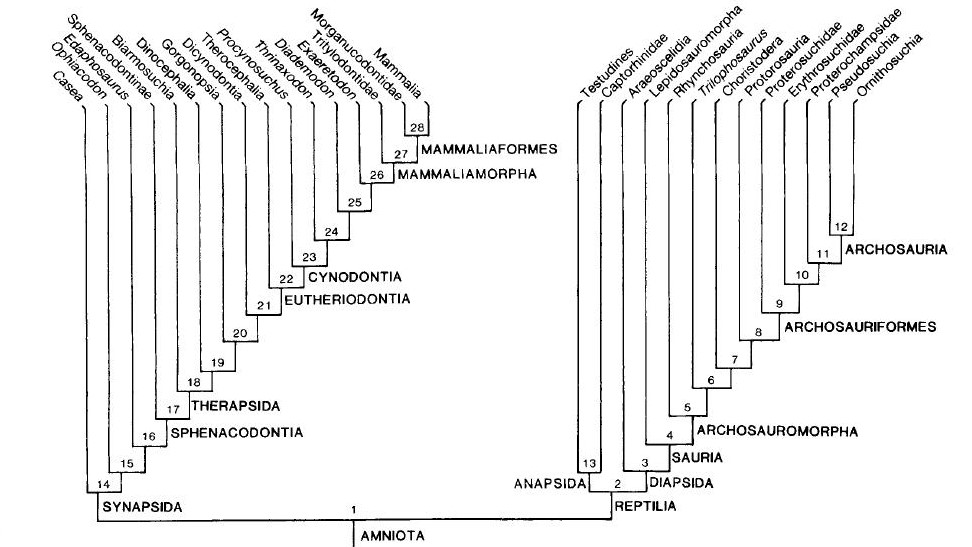
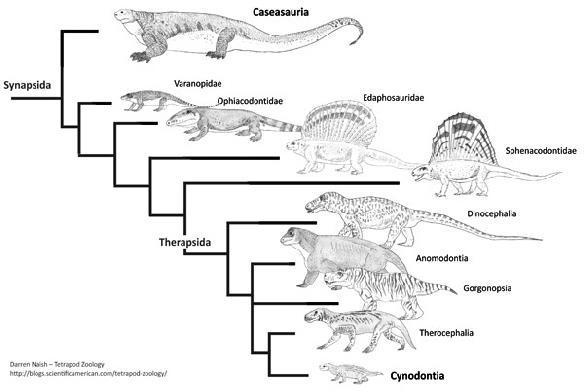
De acordo com os autores, quanto mais antiga é a origem de uma linhagem e quanto mais derivada for a morfologia de seus representantes atuais (como é o caso dos mamíferos), mais os fósseis serão importantes para elucidar sua evolução. Basta compararmos os mamíferos atuais com seus parentes extintos, os “pelicossauros” como Casea, Ophiacodon, Edaphosaurus, Sphenacodon (figura 4) para ver o “abismo morfológico” que separam essas linhagens. Quando observamos os mamíferos atuais, dificilmente conseguimos relaciona-los com outro grupo de animal vivente e quando tentamos, certamente estaremos fadados ao erro, assim como Gardiner, em 1982 equivocadamente agrupou Aves e Mammalia no antigo grupo Haeomothermia. Porém, quando olhamos para os fósseis vislumbramos um mundo completamente “novo”. As formas extintas possuem combinações únicas de características basais e derivadas que frequentemente se apresentam numa serie gradual de mudanças, que nos permitem acompanhar suas histórias evolutivas de maneira tal que possamos observar espécies diferentes ficando cada vez mais semelhantes conforme voltamos no tempo. Os fósseis são uma das poucas evidências diretas da evolução e é por isso, que os dados paleontológicos se tornam tão fundamentais para elucidar as relações evolutivas dos grupos viventes. Principalmente, quando o objeto de estudo são animais tão diferentes como seu cachorro e um pardal na janela de casa.
Outro experimento que a equipe de Gauthier conduziu consistiu basicamente em retirar todas as linhagens viventes da matriz de características e fazer uma nova análise. Apesar de ser um experimento simples, o resultado obtido foi revelador: a nova hipótese representada na figura 3 veio à tona novamente, porém sem os animais viventes. Este resultado, associado à análise inicial das características utilizadas no trabalho de Gardiner (1982), derruba os argumentos dos críticos a respeito da incompletude do registro fóssil. De fato a informação que provem dos fosseis é mais incompleta do que as que podemos retirar dos animais viventes, mas mesmo assim os fósseis estão longe de serem menos informativos em uma análise filogenética. Gauthier demonstrou isso quando retirou todas as espécies viventes da análise e mesmo assim alcançou a nova hipótese. Além disso, Gauthier enfatizou que incompletude não é exclusiva de fósseis. Ela pode ocorrer naturalmente nos animais. Quatro das características utilizadas na análise de Gardiner eram do osso quadrado, um osso na base do crânio dos vertebrados no qual se articula a mandíbula. Contudo, nos sinapsídeos mais derivados, os cinodontes (o qual mamíferos fazem parte), esse osso foi gradativamente sendo reduzido e alocado cada vez mais para trás do crânio junto com uma série de ossos da mandíbula. Esta condição alcançou o extremo nos mamíferos, onde o osso quadrado e um grupo de ossos que uma vez pertenceram à mandíbula, se reduziram e modificaram sua função ao ponto de formarem o que hoje são nossos ossículos do ouvido médio (o quadrado dos outros vertebrados é a bigorna nos mamíferos). Logo, aquelas quatro características baseadas na morfologia do osso quadrado, não se aplicam aos mamíferos, pois o quadrado deles se alterou tanto que fica difícil comparar com os outros animais, ou seja, este é um dado naturalmente faltante. Além disso, Gauthier percebeu que Casea (o fóssil mais basal e antigo de sinapsídeo e que consequentemente esperávamos maior incompletude) apresentava 26% de informação faltando, enquanto que os mamíferos atuais tinham em média 15% de dados morfológicos faltando ou difíceis de interpretar. Esses 11% de diferenças são realmente significantes? Gauthier não só provou que não há diferença significante como também nos mostrou que informação incompleta não é exclusiva dos fósseis.
A Renascença dos fósseis
Após demonstrar empiricamente quão importante os fósseis são para nossa compreensão da evolução das linhagens viventes, Gauthier deu um solavanco nos sistematas, convidando-os a se debruçarem no assunto. Desde então, estudos importantes vêm sendo conduzidos, cada vez mais enaltecendo o uso de dados paleontológicos em reconstruções filogenéticas e desenvolvendo metodologias para minimizar o efeito da incompletude dos dados de organismos tanto viventes quanto extintos, o famigerado missing data que os cladistas tanto abominam (veja Donoghue et al., 1989; Smith, 1998; Wilkinson; Benton, 1995).
Exemplos como estes de Gardiner e Gauthier nos mostram como a Ciência é dinâmica. O que antes era tido como verdade e que hoje é obsoleto, não necessariamente deixa de ser útil. Se Gardiner não tivesse se aventurado nas relações de Amniota, Gauthier e toda uma geração de sistematas não seriam impulsionadas a refletir sobre o assunto e talvez o reconhecimento da importância dos fósseis na reconstrução da evolução da vida na Terra poderia ser ainda mais postergada. Por fim, quando ignoramos as informações contidas no registro fóssil, estamos ferindo o princípio da Evidência Total (um tema que será abordado em postagens futuras do blog). Este princípio pode ser exemplificado com uma ótima analogia que um estimado amigo uma vez me fez: não usar os dados paleontológicos para inferir filogenia é como ter um bebê de colo e joga-lo pela janela só porque ele não é um adulto.
Referências:
AX, Peter. 1987. The phylogenetic system: the systematization of organisms on the basis of their phylogenesis.
DARWIN, C. A Origem das Espécies. Hemus – Livraria Editora Ltda, São Paulo, SP.
DONOGHUE, Michael J. 1989. Phylogenies and the analysis of evolutionary sequences, with examples from seed plants. Evolution, v. 43, n. 6, p. 1137-1156.
GARDINER, BRIAN G. 1982. Tetrapod classification. Zoological Journal of the Linnean Society, v. 74, n. 3, p. 207-232.
GAUTHIER, Jacques; KLUGE, Arnold G.; ROWE, Timothy. 1988. Amniote phylogeny and the importance of fossils. Cladistics, v. 4, n. 2, p. 105-209.
HENNIG, Willi. 1950. Grundzuge einer Theorie der phylogenetischen Systematik.
LOVTRUP, Soren. 1985. On the classification of the taxon Tetrapoda. Systematic Zoology, v. 34, n. 4, p. 463-470.
PATTERSON, Colin. 1981. Significance of fossils in determining evolutionary relationships. Annual Review of Ecology and Systematics, v. 12, n. 1, p. 195-223.
SMITH, Andrew B. 1998. What does palaeontology contribute to systematics in a molecular world?. Molecular phylogenetics and evolution, v. 9, n. 3, p. 437-447.
WILKINSON, Mark; BENTON, Michael J. 1995. Missing data and rhynchosaur phylogeny. Historical Biology, v. 10, n. 2, p. 137-150.

Geovane Alves de Souza, Graduado em licenciatura e bacharelado em Ciências Biológicas pela Universidade Estadual de Londrina. Atualmente é mestrando em Zoologia pelo Museu Nacional/UFRJ. Já desenvolveu pesquisas na área de parasitologia de animais silvestres, hoje conduz estudos de osteohistologia com titanossauros.
















{kind=link}