Este breve artigo procura dar continuidade à história que começamos a contar neste post aqui. A idéia era discorrer sobre a biografia de alguns importantes grupos de dinossauros terópodes, que ajudaram a lançar luz sobre o estudo da evolução das Aves. Para compreender a evolução desses animais, primeiro nós precisamos investigar e aprender mais sobre os principais grupos de dinossauros proximamente relacionados a elas. O foco da narrativa, então, serão os dinossauros coelurossauros.
Os Coelurosauria são um ramo basal dentro do agrupamento dos dinossauros terópodes (dinossauros bípedes que incluem todos aqueles predadores e algumas curiosas famílias de hábitos herbívoros ou onívoros). O nome refere-se à natureza ‘oca’ das vértebras desses animais, e significa, literalmente, “lagarto oco”. Foi dentro deste clado que ocorreu um dos mais fantásticos eventos evolucionários da história dos vertebrados terrestres: A evolução das aves. Ainda que haja alguma controvérsia sobre qual família dinossauriana teria dado origem às aves modernas, é bem consolidado que ela teria pertencido a esse grupo.
No primeiro post sobre “A Origem das Aves” discorremos brevemente sobre a linhagem dos poderosos tiranossauros (aqui), que formariam o grupo mais basal dentro de Coelurosauria. Já no post de hoje, abordaremos um pouco sobre a história dos Ornithomimosauria, os dinossauros que “imitavam aves”.
Figura 1. Cladograma evidenciando as relações evolutivas entre os vários grupos de Coelurosauria. Por Thomas Holz.
Em meados da década de 1980, Jacques Gauthier, então paleontólogo de vertebrados e sistemata na Universidade da Califórnia, Berkeley, identificou em um grupo de dinossauros predadores (terópodes) uma característica que até então era considerada como exclusiva das aves: Penas! Gauthier, entusiasmado, logo inseriu esse novo e importante dado em suas análises filogenéticas. Foi a partir de então, que seu trabalho começou a consolidar àquela que seria conhecida como a hipótese ave-dinossauro. Esta hipótese logo obscureceu as outras menos robustas sobre a origem das aves. Hoje, podemos dizer que a evolução das aves a partir dos dinossauros é uma das mais aceitas e incontestáveis hipóteses dentro do estudos sobre a evolução dos vertebrados. Há um forte apoio dado por um conjunto de caracteres anatômicos peculiares encontrados somente em aves e alguns dinossauros terópodes. Estas características definem não apenas o padrão de evolução do esqueleto entre as aves e dinossauros, mas também abrangem um conjunto de características fisiológicas semelhantes (isto é passível de teste em aves e répteis modernos), que revelam as etapas de transformação de um dinossauro corredor até um pássaro capaz de vôo ativo.
Ornithomimosauria: “Os lagartos que imitavam aves”
Figura 2. Ilustração de Peter Schouten – Grupo de Ornithomimosauria
“De fato, as comparações entre este dinossauro e um avestruz, em vários aspectos de sua anatomia, são tão impressionantes que ele é freqüentemente referido como “o dinossauro mímico de avestruz””.
Edwin Colbert, 1969, em “Evolution of the Vertebrates“, página 201.
Os ornitomimossauros são melhor entendidos quando os imaginamos como uma “versão esportiva” dos dinossauros. Construídos para velocidade, eles eram um grupo de terópodes cuja vida deveria ser muito semelhante àquela das aves ratitas atuais (como os avestruzes, emas e casuares). Eles deveriam, em sua maioria, ser onívoros (apesar de um deles ter sido encontrado com seu conteúdo estomacal preservado indicando uma dieta herbívora – Sinornithomimus) e essencialmente sociais.
Recentemente, a descoberta de membros bastante primitivos desse grupo ajudou a compreender melhor como se deu a evolução e radiação desses animais. Hoje eles são considerados basais dentro da linhagem dos terópodes, e estão colocados entre celurossauros primitivos e o grupo mais derivado destes, que inclui os chamados “maniraptores” (e.g. terezinossauros, alvarezssauros, ovirraptorossauros, tro
odontídeos, dromeossauros e aves) (Figura 1).
O primeiro fóssil de ornitomimossauro foi encontrado em 1889 por George Cannon, em rochas de idade cretácica no Estado do Colorado, Estados Unidos. Charles Othniel Marsh nomeou os fragmentos encontrados – pertencentes à membros posteriores de um animal adulto– como Ornithomimus, significando “o mímico de ave”, já que os considerou muito semelhantes à ossos dos pés de aves atuais.
Figura 3. Gallimimus – Ilustração de Peter Schouten.
Foi somente em 1917, no entanto, que um esqueleto relativamente completo desses animais foi encontrado e a sua anatomia pode ser melhor investigada. Lawrence Lambe foi quem encontrou os restos parciais expostos nos afloramentos de Alberta, mas foi Henry Fairfield Osborn que os nomeou como Struthiomimus, “o mímico de avestruz”. Osborn perceptivamente observou que este dinossauro possuía muitas similaridades à ave ratita atual.
A mais significante série de descobertas de ornitomimossauros, todavia, veio somente a partir da década de 1960, com as expedições poloneso-mongóis e na década de 1990 por meio de sítios recém descobertos na Espanha e na China.
Tudo indicava que estes animais eram construídos para correr rápido. Tratavam-se de velocistas. Possuíam uma cauda longa, horizontal e rígida, pernas alongadas e uma constituição leve, com cabeça pequena e pescoço grácil e alongado. Trilhas de pegadas fossilizadas atribuídas à ornitomimossauros sugerem que estes animais poderiam alcançar velocidades acima de 57km/h, tornando-os os mais rápidos de todos os dinossauros conhecidos, tão velozes quanto cavalos de corrida.
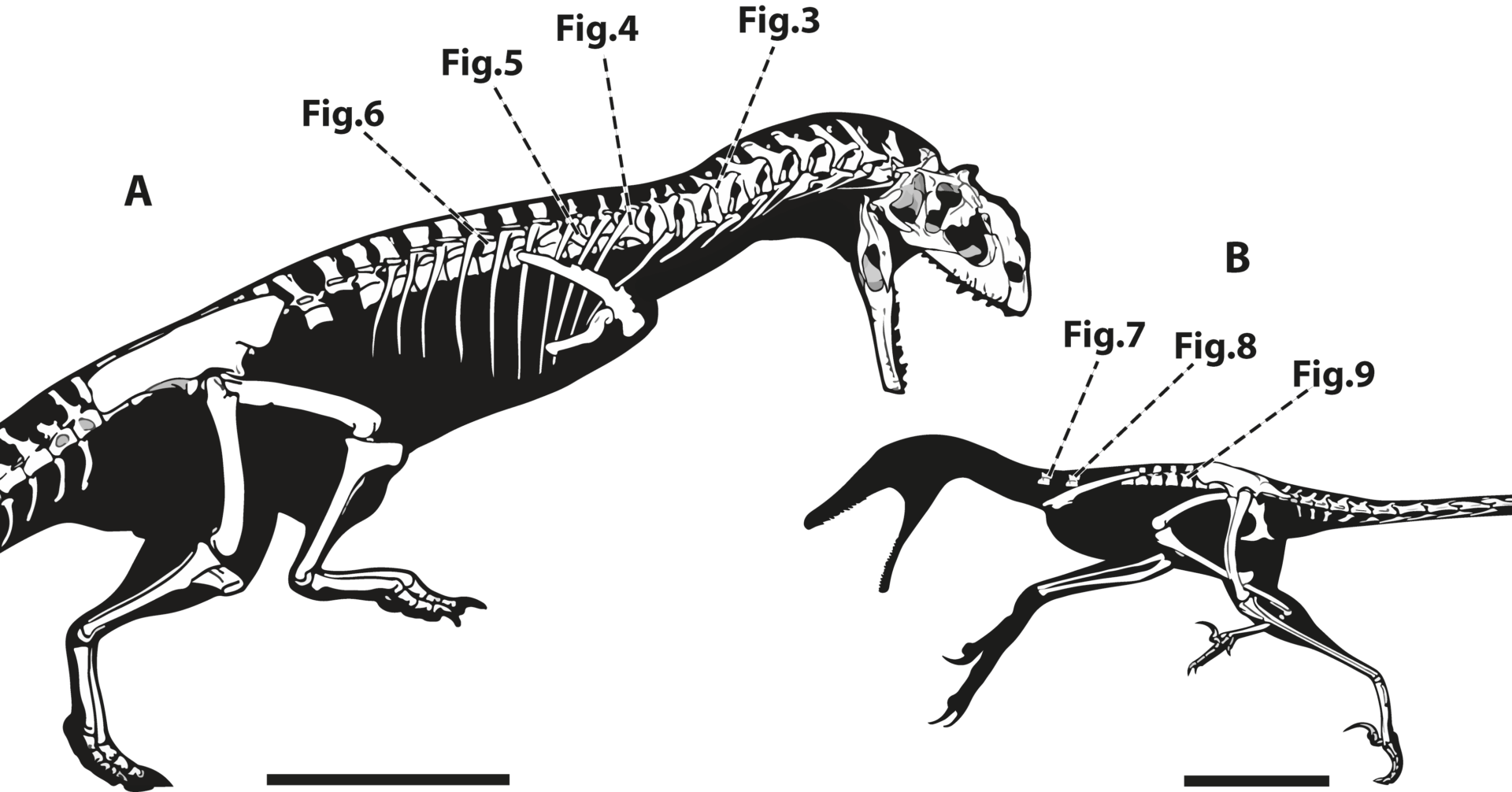
Figura 4. Esqueleto quase completo de ornitomimídeo.
O membro mais primitivo desse grupo é Pelecanimimus, encontrado no famoso depósito de Las Hoyas, Espanha, do início do Cretáceo. Pelecanimimus (“mímico de pelicano”) é, por enquanto, o único membro deste grupo a apresentar numerosos e pequeninos dentes sem serrilha em suas mandíbulas (cerca de 220 no total). Apenas poucos outros ornitomimossauros também possuem dentes como característica, e estes em geral os possuem restritos à parte frontal do rostro somente (e.g. como Shenzhosaurus de Shenzhou, província de Lioning, na China, e Harpymimus de Dundgov, na Mongólia). O restante dos ornitomimossauros não possuem dentes, mas sim bicos bem desenvolvidos. Algumas formas (Shenzhosaurus e Sinornithomimus por exemplo) foram encontradas com pequenos pedregulhos, denominados gastrólitos, em seus estômagos. Estes pedregulhos eram provavelmente utilizados para ajudar a digerir o alimento. Atualmente somente animais herbívoros utilizam-se dessas pedras para auxiliar a moer o material vegetal resistente (sementes, por exemplo) em seus tratos digestivos.

Figura 5. Reconstituição de Pelicanimimus (nome do autor não informado).
Uma teoria postula que os ornitomimossauros deveriam alimentar-se como os flamingos, filtrando a água com os seus bicos curvos, porém outros especialistas discordam, afirmando que as estruturas especiais no palato desses animais eram somente para sustentar o bico e não uma especialização para alimentar-se por filtração.
Não se sabe ainda ao certo como os ornitomimossauros protegiam os seus jovens e filhotes e como organizavam seus bandos. No entanto, recentemente, um conjunto de 14 Sinornithomimus foram encontrados fossilizados juntos na Mongólia. Onze dos animais demonstraram ser jovens ou sub-adultos e estes estavam acompanhados de animais maduros. Os últimos provavelmente estavam protegendo o grupo de mais novos.
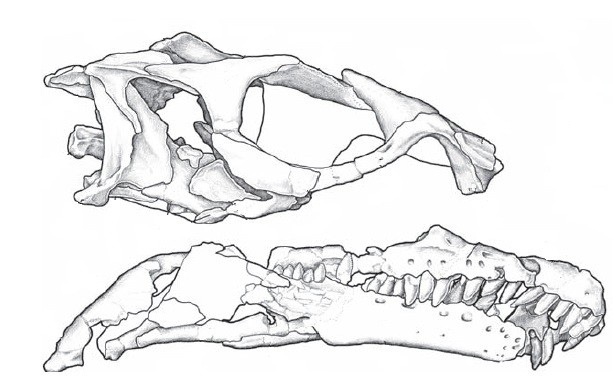
Figura 6. Crânio de Sinornitomimus.
Os ornitomimossauros mais especializados estão divididos em dois grupos. Um norte-americano, que compreende Struthiomimus, Dromiceiomimus e Ornithomimus, e um asiático, composto por Gallimimus e Anserimimus. Ornithomimus demonstra o típico formato de um crânio avançado de Ornithomimosauria, possuindo olhos grandes e um focinho delgado e curvado, com a mandíbula inferior pouco mais alongada que a superior.
As grandes órbitas dos ornitomimossauros são freqüentemente preservadas com os ossículos escleróticos, que ajudavam a sustentar o olho em vida. Tudo indica que esses animais teriam um senso de visão bastante desenvolvido. Eles possuíam um cérebro grande, comparável àquele de aves modernas e inclusive àquele bem desenvolvido dos troodontídeos. Embora o grupo seja caracterizado pela presença de pernas e braços longos, a mecânica dos membros anteriores de Struthiomimus sugere que eles não eram equipados para escavar, mas eram sim instrumentos gráceis mais adequados para agarrar galhos e folhas.
Figura 7. Crânio de Gallimimus.
A herbivoria parece ter evoluído bastante cedo na história evolutiva desse grupo. Shenzhousaurus foi encontrado com gastrólitos em sua região estomacal e esta espécie é considerada como uma forma relativamente primitiva dentro do grupo. Ela é considerada primitiva não somente pela presença de dentes, mas por outras características esqueletais como o comprimento do ‘polegar’, mais curto que os outros dígitos (nas formas derivadas ele tem o comprimento semelhante ao dos outros dedos).
Figura 8. Cladograma de Ornithomimosauria indicando a posição basal de Pelicanimimus e Shenzhousaurus (Makovicky, 2009).
Figura 9. Outro cladograma de Ornithomimosauria. Fonte: www.dinodata.org, baseado em Makovicky et al., 2004.
Uma das mais recentes descobertas do grupo foi Beishanlong, do Cretáceo chinês, animal muito parecido com Harpymimus, outro ornitomimossauro da China. Porém, a característica mais impressionante de Beishanlong é o seu tamanho. Definitivamente trata-se do maior ornitomimossauro já descrito e o mais interessante: análises histológicas demonstraram que o indivíduo analisado ainda encontrava-se em fase de crescimento. A partir do material encontrado foi estimado para o animal uma massa de 626kg. É interessante observar um paralelismo do gigantismo em diferentes linhagens de celurossauros herbívoros (ver Shuzhousaurus (Therezinosauria) e Gigantoraptor (Oviraptorosauria)).
Figura 10. Esquema indicando o tamanho de Beishanlong proporcionalmente a um ser humano.
Inicialmente acreditava-se que estes animais estariam presentes somente na América do Norte e na Ásia, mas depois da descoberta do primitivo Pellicanimimus na Europa, a história começou a ficar mais clara e as noções biogeográficas e de origem do grupo a solucionarem-se. Sem dúvida, o que pode-se dizer por enquanto, é que trata-se de um grupo restrito aos continentes do norte (laurasianos), com sua origem provavelmente européia.
Apesar das tantas semelhanças com as aves, que foram inclusive batizados em alusão a elas, não foi dentro deste grupo que as nossas colegas emplumadas tiveram sua origem. Os Ornithomimosauria são apenas parentes próximos. A história continua. Sem dúvida as aves tiveram origem dentro do grupo dos Maniraptora e é para aí que se aproxima ainda mais a próxima parte de nossa história.
No próximo capítulo: À um passo dos maniraptora…
Referências Bibliográficas:
Makovicky, P. J., Kobayashi, Y. & Currie, P. J. 2004. Ornithomimosauria. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds) The Dinosauria, Second Edition. University of California Press (Berkeley), pp. 137-150.
Barrett, P. M. 2005. The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria). Palaeontology48, 347-358.
Kobayashi, Y. & Lü, J.-C. 2003. A new ornithomimid dinosaur with gregarious habits from the Late Cretaceous of China. Acta Palaeontologica Polonica 48, 235-259.
Norell, M. A., Makovicky, P. & Currie, P. J. 2001. The beaks of ostrich dinosaurs. Nature 412, 873-874.
Russell, D. A. 1972. Ostrich dinosaurs from the Late Cretaceous of western Canada. Canadian Journal of Earth Sciences 9, 375-402.
Long, J. & Schouten, P., 2008. Feathered Dinosaurs: The Origin of Birds. Oxford university Press. Oxford, New York.
Naish, D. – The Tet Zoo field guide to ostrich dinosaurs (Part I) and (Part II). In: Tetrapod Zoology.


























{kind=link}
{kind=link}