Hoje (quarta-feira 9/12/2020), acaba de ser publicado um trabalho que traz informações importantes para a compreensão de um grande enigma da paleontologia: a origem dos pterossauros, répteis voadores que dominaram os céus durante a Era Mesozoica.
O trabalho foi publicado na revista Nature e conta com a participação de alguns cientistas brasileiros, que ajudaram a investigar em detalhes um grupo de pequenos répteis que viveu durante o Período Triássico, os lagerpetídeos.

A origem dos pterossauros sempre foi um daqueles problemas cabeludos da Paleontologia. Os pterossauros, para quem não conhece ou não se recorda, são aqueles répteis voadores com asas membranosas, que viveram durante a chamada “Era dos Dinossauros”, intervalo do tempo que se estende de 251 à 66 milhões de anos atrás.
O problema com os pterossauros é que seus fósseis mais antigos (que tem cerca de 208 milhões de anos), já apresentam características tão diferenciadas, que é difícil traçar a origem evolutiva do grupo. Eles já apresentam, por exemplo, dedos das mãos hiper-alongados, crânio super modificado e diversas outras características derivadas muito bem adaptadas para o voo. A comparação com outros grupos de répteis que viveram no mesmo período é bastante difícil, pois não temos registros de fósseis de organismos com características intermediárias. Logo, descobrir onde o grupo se encaixa na árvore da vida tornou-se um desafio.
Diversos grupos independentes de cientistas têm se debruçado sobre a questão ao longo do tempo. Os resultados específicos são bastante contrastantes, mas em uma coisa, pelo menos, temos concordado, eles pertencem a um grupo chamado de Archosauromorpha, que inclui, por exemplo, os dinossauros e os crocodilos atuais. É um grupo muito amplo. Isso serve para você entender a gravidade da questão.
Há muita discordância sobre em que ponto exato dentro dos Archosauromorpha se encaixam os pterossauros. Alguns pesquisadores sugerem que eles teriam derivado de Archosauromorpha basais. Outros, que eles são parentes bem mais próximos dos dinossauros, derivados de avemetatarsálios basais próximo aos lagerpetídeos. Aí entra o estudo publicado hoje.

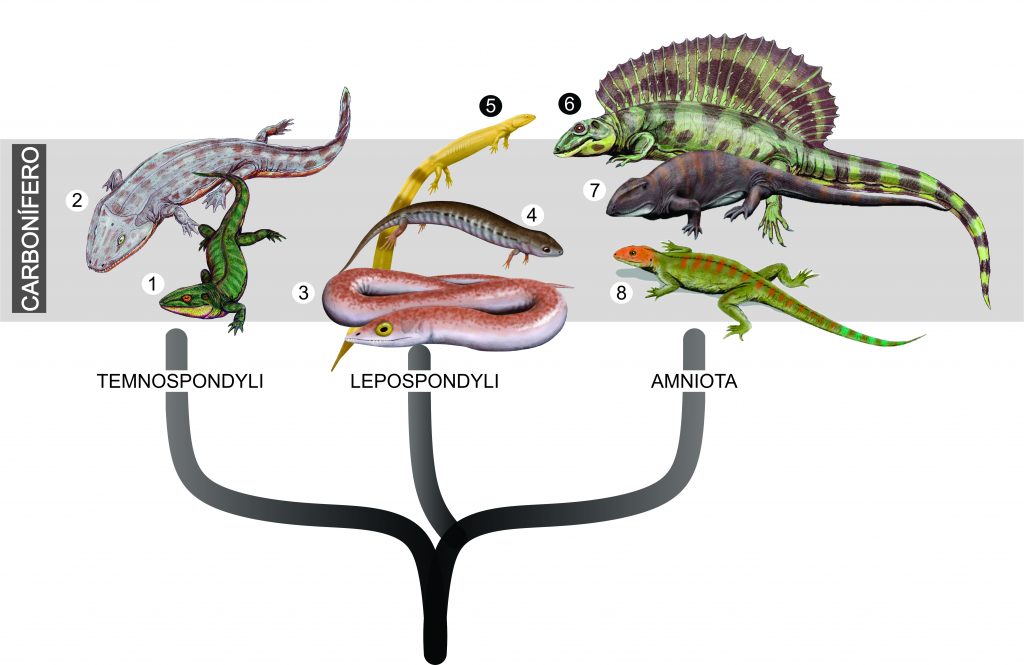
O estudo publicado hoje na revista Nature é assinado por 18 autores, entre eles os brasileiros Mário Bronzati, Sergio F. Cabreira, Lúcio Roberto da Silva e Max Langer. O que os 18 pesquisadores fizeram foi investigar mais a fundo um grupo específico de pequenos répteis arcossauromorfos do Período Triássico (cerca de 237 a 210 milhões de anos atrás), conhecidos como lagerpetídeos. A equipe analisou não apenas detalhes da aparência externa do esqueleto desses organismos, como utilizou também a tecnologia de tomografia computadorizada para observar dentro de seus ossos. Mais especificamente, dentro do crânio.
Os lagerpetídeos viveram onde hoje é a América do Sul, América do Norte, África e Madagascar. Durante o Triássico, estas massas de terra estavam unidas, formando o supercontinente Pangea. Aqui no Brasil, fósseis de lagerpetídeos são encontrados apenas no Rio Grande do Sul, em rochas da Bacia do Paraná.
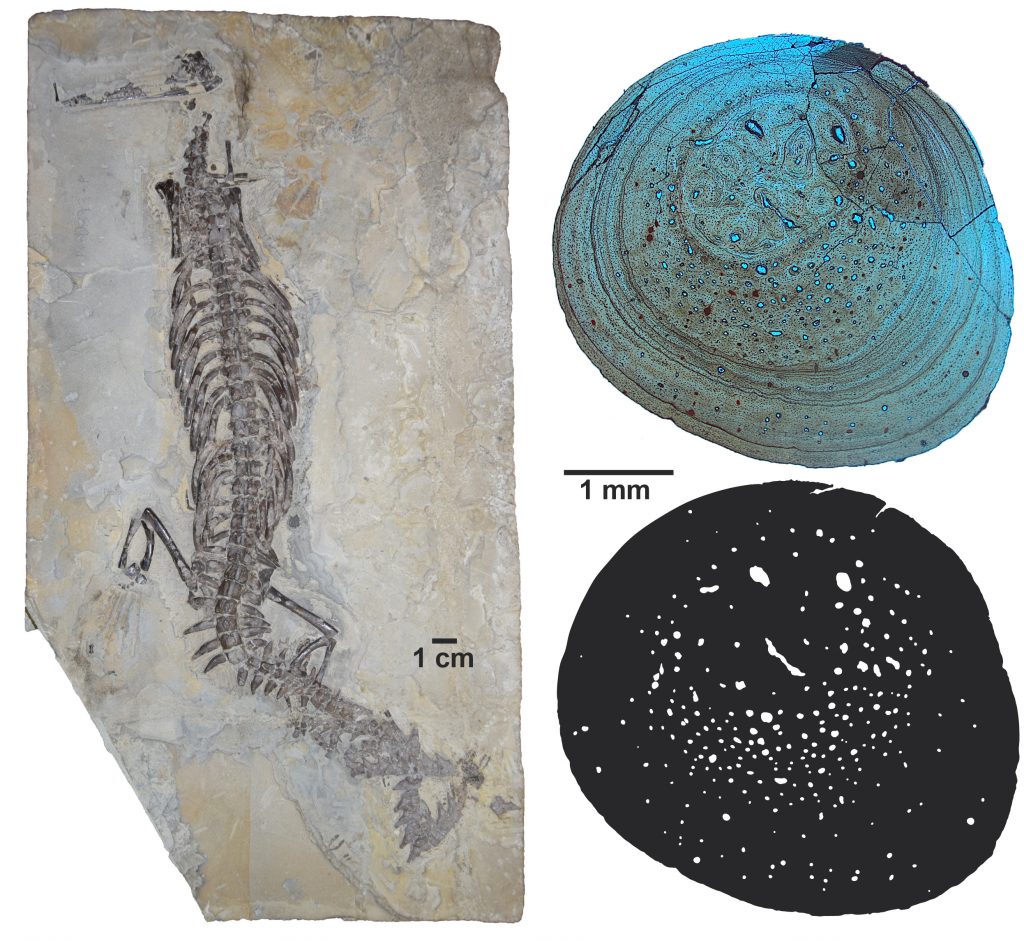
Um exemplo de lagerpetídeo brasileiro é Ixalerpeton polesinensis, um animal com cerca de 40 cm de comprimento e 15 centímetros de altura, descrito em 2016. Seu fóssil foi encontrado no município de São João do Polêsine, RS. Ixalerpeton é uma das espécies de lagerpetídeos incluída no estudo. Ele tem preservado parte de sua caixa craniana, o que permitiu aos pesquisadores reconstruir o cérebro e ouvido interno do animal.

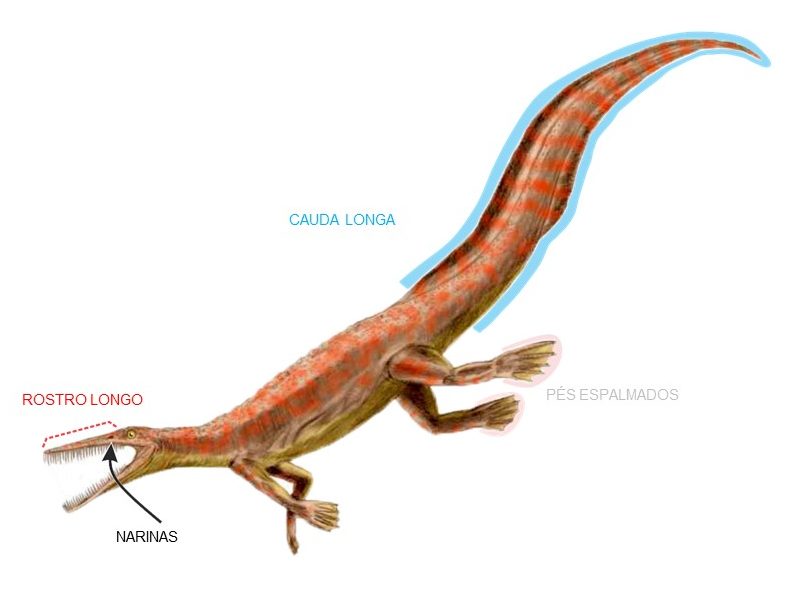
Outras espécies de lagerpetídeos foram analizadas, como Lagerpeton chanarensis, da Argentina, Dromomeron romeri e Dromomeron gregorii, dos E.U.A. e Kongonaphon kely, de Madagascar. Até pouco tempo atrás, lagerpetídeos eram conhecidos apenas com base em ossos das pernas e do quadril (que se preservam mais facilmente nesse grupo), mas descobertas mais recentes têm revelado mais detalhes sobre a anatomia desses animais, como a aparência do crânio, coluna e membros anteriores.
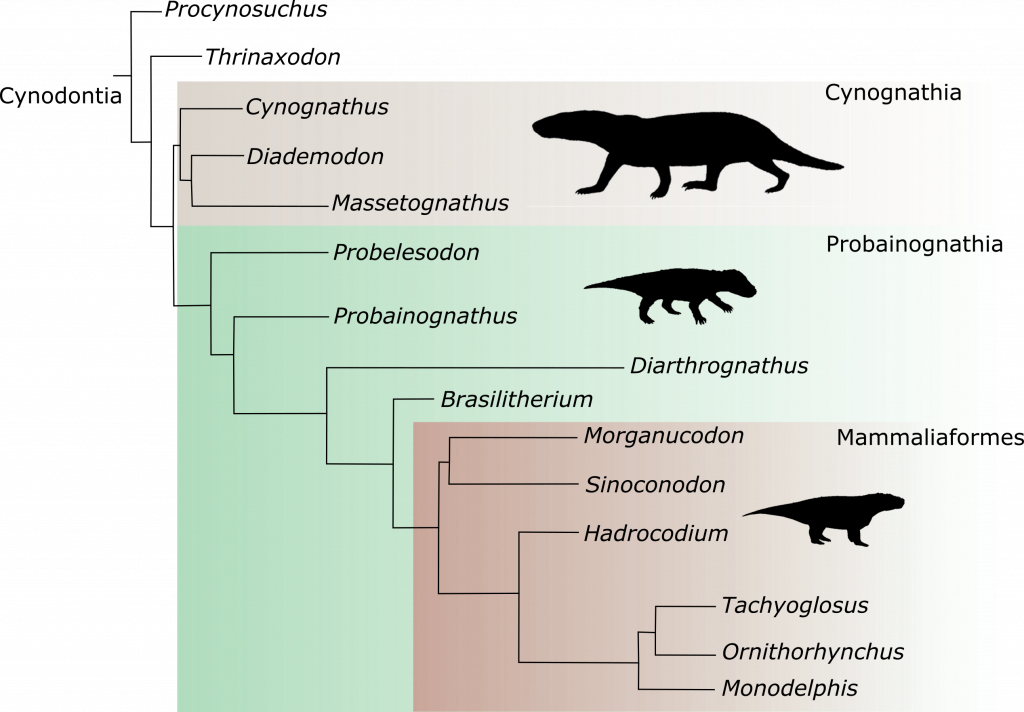
Os pesquisadores reuniram, então, esses fósseis e fizeram um estudo anatômico comparativo, avaliando diversas características destes animais e comparando-as com de outros Arcosauromorpha. A análise filogenética resultante do estudo (uma forma matemática de análise de parentesco) revelou grandes semelhanças anatômicas entre os lagerpetídeos e os primeiros pterossauros. Apontou, inclusive, a existência de um grupo independente, que incluiria ambas as linhagens, chamado de Pterosauromorpha.

No trabalho são destacadas várias características únicas compartilhadas apenas por pterossauros e lagerpetídeos, incluindo, por exemplo, alongamento dos ossos do antebraço e das mãos. O parentesco muito próximo surpreendeu bastante, pois anteriormente pensava-se que lagerpetídeos deveriam ser mais aparentados aos silesaurídeos e dinossauros. Assim, o atual estudo estabelece um marco importante para o entendimento da origem dos pterossauros.
Outro resultado fascinante do estudo publicado hoje, envolve a biologia dos lagerpetídeos. Algumas características anatômicas indicam adaptações a um estilo de vida bastante ágil. As análises com tomografia computadorizada e reconstrução do cérebro e ouvido interno desses animais, por exemplo, demonstraram que eles tinham sistemas sensoriais típicos de organismos capazes de movimentos rápidos da cabeça e bom controle do movimento dos olhos e do pescoço. Adaptações como essa são observadas em organismos voadores e/ou arborícolas da atualidade. Talvez os lagerpetídeos utilizassem essa sua característica para a captura de presas ágeis, como insetos. O que é reforçado pela anatomia dos seus dentes.

Se comparado ao sistema sensorial dos pterossauros, lagerpetídeos tinham basicamente as mesmas adaptações no cérebro e ouvido interno. Isso pode sugerir que características sensoriais vantajosas para o voo evoluíram antes mesmo do voo em si nos Pterosauromorpha (uma pré-adaptação).
Outro ponto interessante é o formato das garras das mãos dos lagerpetídeos, bastante curvadas, o que indica uma função diferenciada dos membros anteriores. Os autores sugerem que elas podem ter sido selecionadas devido a um estilo de vida arborícola (para ajudar a escalar) e/ou utilizadas para a aquisição de presas. O que é interessante , pois pode sugerir um cenário em que o voo nos pterossauros teria evoluído como uma vantagem para se mover de uma árvore para a outra.

Embora este estudo tenha demonstrado um parentesco entre lagerpetídeos e pterossauros, muitas questões ainda permanecem. Entre elas, talvez a mais perturbadora ainda seja como a principal característica dos pterossauros, as asas, evoluíram. Pode parecer frustrante não ter todas as respostas de uma vez, eu sei. Mas estamos chegando cada vez mais perto. Este estudo nos aproxima um pouco mais da “solução” do grande enigma. Pense pelo lado bom: pelo menos, agora sabemos melhor onde procurar respostas!
Nesta sexta-feira, dia 11/12, às 19h, vamos bater um papo ao vivo no nosso canal do Youtube com um dos autores do estudo. Ative o lembrete para não perder:
Veja o vídeo de divulgação do artigo:
Referência:
Ezcurra, M. D. et al. 2020. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature. https://www.nature.com/articles/s41586-020-3011-4
Leia também:
Padian, K. 2020. Closest relatives found for pterosaurs, the first flying vertebrates. Nature https://www.nature.com/articles/d41586-020-03420-z