Molécula biológica responsável pela pigmentação de seres vivos foi encontrada preservada em um fóssil brasileiro de cerca de 110 milhões de anos, da região do Ceará! O fóssil em questão é de um pterossauro, um tipo de réptil voador da “Era dos Dinossauros”.
Reconstituição em vida de Tupandactylus, arte de Márcio Castro.
O estudo foi publicado hoje em uma das revistas científicas do prestigioso grupo Nature, a Scientific Reports, e inclui pesquisadores diversos países, liderados pelos paleontólogos brasileiros Felipe Pinheiro, da Universidade Federal do Pampa (Unipampa, Rio Grande do Sul) e o doutorando Gustavo Prado, da Universidade de São Paulo (USP, São Paulo).
“Isso ainda é muito distante Jurassic Park”, lembram os pesquisadores, mas o fato de encontrar uma molécula biológica tão bem preservada já é uma grande descoberta, que nos possibilita entender melhor como eram esses organismos do passado.
O fóssil de réptil voador analisado pertence a um Tupandactylus, um pterossauro de tamanho médio, com cerca de 3 metros de envergadura e que tinha uma crista bem alta na cabeça. Ele viveu no sul do Ceará, na região do Araripe, quando toda essa área era coberta por uma extensa laguna, durante a primeira metade do Período Cretáceo, há cerca de 110 milhões de anos.
O estudo também contou com a participação de pesquisadores do Japão e dos Estados Unidos. Trata-se da mais completa caracterização química de uma biomolécula fossilizada em um réptil.
“Embora sempre soubéssemos que os fósseis encontrados na região da Chapada do Araripe eram especiais em termos de preservação, foi uma surpresa quando as análises químicas mostraram que a melanina do bicho ainda estava lá. Parece que o pterossauro morreu ontem”, relata Felipe Pinheiro, paleontólogo da Unipampa.
Vários fósseis de Tupandactylus já foram descobertos na Chapada do Araripe. Porém, este preservou muito bem a crista do animal, o que levou os pesquisadores a quererem analisá-la mais de perto. A crista enorme, em forma de vela, provavelmente era utilizada, entre outras coisas, para atrair parceiros. Foi dela que os cientistas extraíram o pigmento.
Imagem do artigo monstrando os pontos amostrados no fóssil.
“A melanina é uma das moléculas mais resistentes aos processos de fossilização. Enquanto os outros compostos são degradados com o passar do tempo, esse pigmento resiste de forma mais ou menos intacta”, explica Gustavo Prado, que é especialista em pigmentos fossilizados.
Imagem do artigo, mostrando os corpúsculos esféricos presentes no fóssil, que conteriam a melanina.
Agora, a pergunta que não quer calar: Com essa molécula preservada, foi possível identificar a cor do animal?
Os cientistas que assinam o estudo são bastante céticos: “É complicado”, diz Pinheiro. “São muitos fatores envolvidos na coloração de um animal, e a melanina é só um deles”. Estudos anteriores reconstruíram a cor de aves e dinossauros com base na forma dos melanossomos, organelas responsáveis por armazenar melanina. A ideia é que o formato dos melanossomos poderia indicar a coloração. A caracterização química da melanina do Tupandactylus mostrou que não é bem assim. “Não encontramos correlação entre o formato dos melanossomos e o tipo de melanina identificada no pterossauro”, diz Gustavo Prado.
O novo estudo, portanto, desafia as inferências de coloração realizadas para organismos fósseis até então. Será necessário rever essa possibilidade e, à luz das novas descobertas, aperfeiçoá-la.
O grupo de pesquisadores continua investigando a preservação excepcional de fósseis da Chapada do Araripe, e afirmam que várias novidades ainda estão por vir. “Aos poucos ficamos cada vez mais próximos desses animais incríveis”, diz Pinheiro.
***
O trabalho completo está disponível em www.nature.com/articles/s41598-019-52318-y
O estudo foi financiado pelo Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e pela Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Estudo publicado nesta quarta-feira (26/06/19) na revista Scientific Reports, do grupo Nature, apresenta uma nova espécie de dinossauro brasileiro, que viveu no Período Cretáceo, há cerca de 90 milhões de anos.
Reconstrução em vida de Vespersaurus paranensis. Crédito da imagem: Rodolfo Nogueira.
O fóssil foi encontrado no município de Cruzeiro do Oeste, PR, e foi estudado por paleontólogos das universidades de São Paulo (USP) e Estadual de Maringá (UEM), além de pesquisadores do Museo Argentino de Ciências Naturales e do Museu de Paleontologia de Cruzeiro do Oeste. A nova espécie foi nomeada Vespersaurus paranaensis.
Vesper (do latim) significa oeste/entardecer, em referência ao nome da cidade onde foi descoberto o fóssil, e paranaensis faz uma homenagem ao Estado do Paraná, já que este é o primeiro dinossauro paranaense descrito.
Os fósseis da nova espécie de dinossauro pertencem a um grupo de dinossauros carnívoros chamados de Noasaurinae. Os Noasaurinae são abelissauros diferentões, de pequeno porte, encontrados desde a Argentina até Madagascar (com possíveis registros na Índia). Estes terópodes viveram em uma época em que os continentes do sul ainda estavam unidos, formando o Gondwana, e transitavam de um lado para o outro, cruzando um imenso deserto que existia entre o Brasil e a África.
Restos de Noasaurinae já eram conhecidos para o Brasil (veja Lindoso et al., 2012 e Brum et al., 2016), mas este é o material mais completo encontrado até o momento. É também o material mais completo de dinossauro terópode descrito para o Brasil até agora, com quase metade do esqueleto encontrado.
Representação tridimensional do esqueleto de Vespersaurus paranensis indicando (em cor sólida) os ossos que foram encontrados. Crédito da imagem: Rodolfo Nogueira.
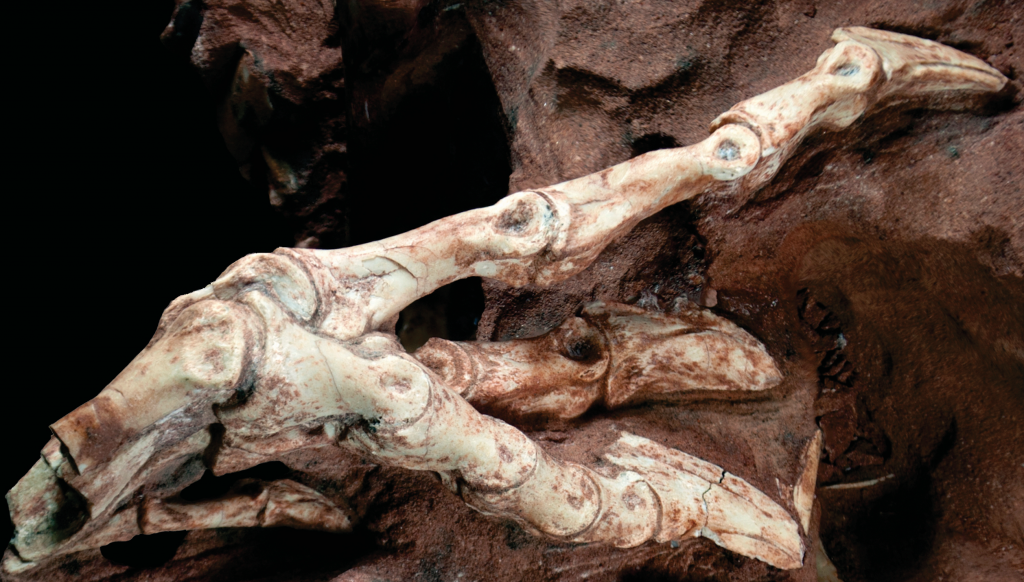
O novo dinossauro possuía vértebras pneumáticas, que conferiam leveza ao seu esqueleto, como nas aves viventes, e um braço muito reduzido (com menos da metade do comprimento da perna). Porém, a sua característica anatômica mais peculiar eram os pés. Seu peso era praticamente todo suportado por um único dedo central, sendo o animal funcionalmente monodáctilo, como os cavalos. Os dedos que flanqueavam o dígito central, por sua vez, possuíam grandes garras em forma de lâmina, que deveriam servir para cortar e raspar carne.
Pata direita de Vespersaurus paranensis como preservada na rocha, note a garra do quarto dedo em forma de lâmina. Foto de Paulo Manzig.
As rochas do noroeste paranaense, nas quais Vespersaurus foi preservado formaram-se em ambientes desérticos, o que sugere que o animal deveria ser adaptado a esse tipo de clima. Na década de 70, em rochas relacionadas, o paleontólogo Giuseppe Leonardi descobriu uma ampla assembleia de pegadas fósseis. Algumas, feitas por um pequeno dinossauro bípede, carnívoro, aparentemente monodáctilo. À época não se conhecia nenhum animal com tais características ao qual elas pudessem ser atribuídas. Muito tempo depois, o produtor parece ter sido encontrado.
Reconstrução em vida do pé de Vespersaurus paranensis. Crédito da imagem: Rodolfo Nogueira.
Vespersaurus paranaensis não é primeira espécie cretácica a ser encontrada no noroeste do Paraná. No mesmo sítio fossilífero em Cruzeiro do Oeste foram descobertos também o lagarto Gueragama sulamericana e inúmeros indivíduos do pterossauro Caiuajara dobruskii. A descoberta de mais uma espécie fóssil em Cruzeiro do Oeste deve impulsionar as pesquisas paleontológicas na região.
Veja o artigo:
Langer et al., 2019. A new desert-dwelling dinosaur (Theropoda, Noasaurinae) from the Cretaceous of south Brazil. Scientific Reports https://www.nature.com/articles/s41598-019-45306-9
Demais referências:
Brum, A.S., Machado, E.B., de Almeida Campos, D. and Kellner, A.W.A., 2016. Morphology and internal structure of two new abelisaurid remains (Theropoda, Dinosauria) from the Adamantina Formation (Turonian–Maastrichtian), Bauru Group, Paraná Basin, Brazil. Cretaceous Research, 60, pp.287-296.
Lindoso, R.M., Medeiros, M.A., de Souza Carvalho, I. and da Silva Marinho, T., 2012. Masiakasaurus-like theropod teeth from the Alcântara Formation, São Luís Basin (Cenomanian), northeastern Brazil. Cretaceous Research, 36, pp.119-124.
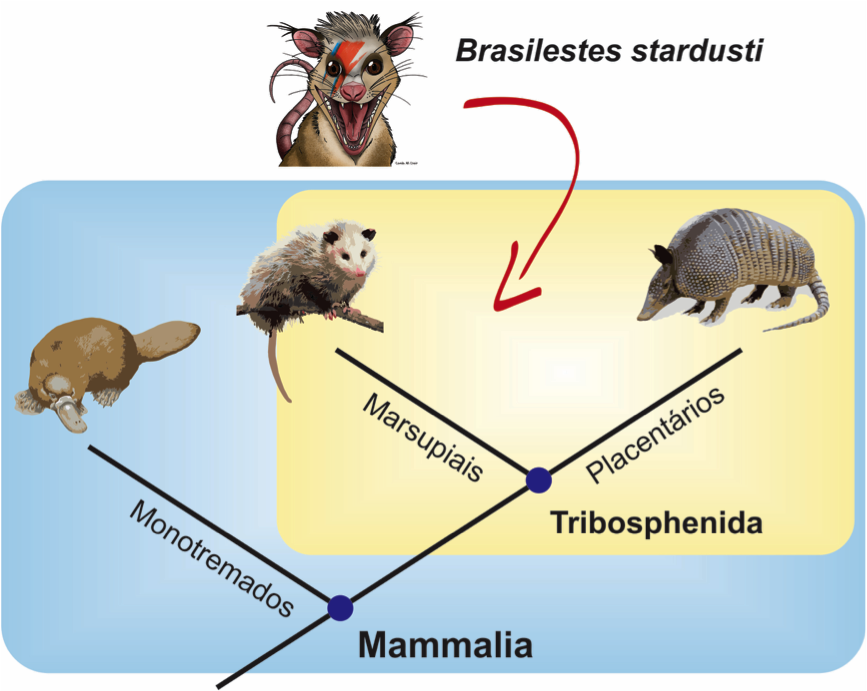
Brasilestes stardusti é o seu nome, em alusão ao Brasil e a Ziggy Stardust, um personagem criado pelo músico britânico David Bowie, falecido em 2016, ano em que o fóssil foi descoberto.
O fóssil em questão tem apenas 3,5mm e trata-se de um único dente pré-molar. Pode não parecer grande coisa, mas é uma descoberta há muito tempo esperada. Tanto que, apesar de não escrever há muito tempo no blog, achei que isto, particularmente, merecia uma comemoração!
A publicação do material foi feita hoje, na revista científica Royal Society Open Science e conta com a participação de pesquisadores brasileiros da UFG, USP e Unicamp, além de paleontólogos argentinos e estadunidenses. O estudo foi liderado pela Dra. Mariela Castro (UFG), especialista em mamíferos fósseis e, com certeza, uma grande fã de Bowie.
O pequeno dente, tão importante, foi encontrado nas rochas ricas em fósseis do interior do estado de São Paulo, mais especificamente, no município de General Salgado, oeste paulista. Estas rochas datam do final Período Cretáceo, entre 80 e 75 milhões de anos, época em que os dinossauros ainda reinavam soberanos nos ecossistemas terrestres. Isso torna Brasilestes o mais antigo mamífero conhecido para o Brasil.
Com um tamanho reconstituído aproximado do de um gambá atual (cerca de 50cm de comprimento), Brasilestes certamente se esquivou das passadas de gigantes pescoçudos herbívoros e fugiu das ferozes mandíbulas de uma miríade de crocodilos terrestres, uma pequena amostra da paleofauna que habitava o interior de São Paulo no final do Cretáceo.
Até a presente data, nenhum vestígio corporal de mamífero fóssil havia sido apropriadamente descrito para as rochas cretácicas do Brasil. Na verdade, para todas as rochas da Era Mesozoica brasileira. O fóssil “do tal mamífero”, sempre havia sido o “Santo Graal” da paleontologia brasileira, buscado incansavelmente por vários grupos de pesquisadores. Por isso, é uma alegria ele ter sido finalmente encontrado.
Para não dizer que este é realmente o primeiro registro corporal de mamífero cretácico do Brasil, o fóssil de um pequeno pedaço de mandíbula com um único dente inserido havia sido encontrado em rochas do mesmo contexto geológico na década 1990. Apesar de publicado há tempos, o material não foi descrito apropriadamente na época, e encontra-se, até hoje, inacessível para a grande maioria dos paleontólogos brasileiros.
Brasilestes é muito importante, porque fornece a primeira identificação mais precisa sobre um mamífero do Cretáceo do Brasil. A morfologia do dente encontrado indica que o mesmo pertencia a um mamífero Tribosphenida, ou seja, um mamífero do grupo que reúne os placentários e marsupiais.
Os mamíferos tribosfênidos contam com diversos registros no Cretáceo do hemisfério norte. Porém, no hemisfério sul, apenas haviam sido encontrados na Índia, em Madagascar e no norte da América do Sul. A descrição de Brasilestes stardusti veio preencher uma lacuna importante no registro de mamíferos mesozoicos, ressaltando a importância do Brasil para a compreensão da história evolutiva do grupo.
Por mais absurdo que possa parecer descrever uma nova espécie com base em um único dente, essa é uma situação muito comum para mamíferos fósseis. O registro fossilífero desse grupo é relativamente raro e, majoritariamente baseado na ocorrência de dentes isolados, lembrando que dentes são elementos muito resistentes e preservam-se com maior facilidade do que ossos. Por esta razão, para mamíferos fósseis, características da dentição são muito utilizadas como caracteres diagnósticos de espécies
A equipe envolvida no trabalho também realizou outras análises no fóssil para ajudar em sua identificação, como a avaliação da microestrutura do esmalte dentário.
A cobertura de esmalte encontrada, com cerca de 20 mícrons, é bem mais fina do que a dos dentes de outros mamíferos mesozoicos (entre 100 e 300 mícrons). Além disso, poucas espécies de mamíferos atuais têm característica semelhante, entre eles, alguns Xenarthra (ordem que inclui os tatus, tamanduás e preguiças). Esta observação suscita uma possível relação entre Brasilestes e este grupo de mamíferos. A divergência (ou origem) dos xenartros, calculada por meio da técnica chamada de “relógio molecular”, teria se de dado exatamente nessa época, há pelo menos 85 milhões de anos.
Os mamíferos eram elementos relativamente raros em ecossistemas da Era Mesozoica. Ocupavam nichos secundários, eram, via de regra, pequenos e predominantemente generalistas. A maioria, talvez tivesse hábitos noturnos e fosse arborícola ou fossorial. Essas são algumas razões para os fósseis de Brasilestes serem tão raros. Provavelmente estes organismos eram entidades pouco abundantes em seu paleoecossistema e/ou tinham tamanhos muito pequenos para que seus elementos ósseos delicados fossem preservados e/ou vivessem longe da área favorável para preservação de fósseis.
Tenho certeza de que esta publicação traz novas esperanças aos paleontólogos brasileiros. Que mais fósseis sejam encontrados em breve e ajudem a responder as questões que Brasilestes trouxe à tona (veja mais aqui). Para fechar, vale sempre a pena lembrar, que os mortos têm muita história para contar. Este, especificamente, marcou seu nome na “calçada da fama” da paleontologia nacional, tal qual o grande artista David Bowie deixou sua marca no famoso passadiço hollywoodiano.
Referência:
Castro MC, Goin FJ, Ortiz-Jaureguizar E, Vieytes EC, Tsukui K, Ramezani J, Batezelli A, Marsola JCA, Langer MC. 2018 A late Cretaceous mammal from Brazil and the first radioisotopic age for the Bauru Group. R. Soc. open sci. 5: 180482. http://dx.doi.org/10.1098/rsos.180482
Olá caros leitores, depois de um breve período de pausa nas postagens, voltamos com grande estilo. Hoje apresento a vocês um interessante texto redigido pelo Mestrando em Zoologia do Museu Nacional/UFRJ Geovane Alves de Souza, O assunto abordado se refere as primeiras discussões históricas sobre as hipóteses filogenéticas dos Amniotas (grupo que tradicionalmente inclui os répteis, aves e mamíferos e suas formas relacionadas) e a importância dos fósseis para um melhor entendimento da evolução das espécies. Então, sem mais delongas, vamos ao texto!
Editado em 19/08/2018.
Em plena Era da Filogenômica, na qual o DNA possui papel central na busca pela compreensão da evolução da vida na Terra, é comum pensarmos nos fósseis como uma fonte de dados um tanto quanto ultrapassada, trabalhosa e que demanda muito tempo para estudar. A diretora do Jurassic World, Claire Dearing (encenada pela atriz Bryce Dallas) enfatiza isso muito bem em sua fala: “Aprendemos mais com a genética em 10 anos do que em um século, escavando”. Hoje é consenso no meio acadêmico a importância dos vestígios da vida pretérita, os fósseis (para melhores detalhes acerca de sua definição veja aqui), quando inferimos relações de parentesco dos organismos viventes. Contudo, nem sempre os fósseis tiveram sua importância reconhecida e passando por momentos de glória e queda ao longo dos últimos séculos.
Após a publicação da obra A Origem das Espécies por Meio da Seleção Natural por Charles Darwin em 1859, no qual o autor defendia as teorias de Evolução Biológica, Seleção Natural e Ancestralidade Comum, os fósseis passaram a desempenhar um papel chave na compreensão de como a vida evoluiu. Contudo, foi na metade do século XX, que o registro fossilífero enfrentou uma queda brusca de sua supremacia. O responsável foi o advento de uma nova maneira de se estudar a evolução: a Sistemática Filogenética de Willi Hennig (1950). A nova metodologia e filosofia da sistemática nos seus primeiros anos de existência não exigia a necessidade de determinar uma dada espécie fóssil conhecida como ancestral entre duas linhagens. Podíamos estudar a evolução das espécies viventes, tratando o ancestral comum entre elas como uma espécie hipotética. Esse modo de vislumbrar as árvores filogenéticas (diagramas ramificados que representam a evolução de uma linhagem e que são gerados a partir de uma análise computacional ou análise filogenética), conhecido como Modelo Cladogenético, é antagônico ao antigo modelo vigente, o Anagenético. De fato, determinar em qual ponto exato da evolução de uma linhagem uma espécie fóssil esta inserida não é uma tarefa fácil, eu diria que um tanto quanto impossível, a menos que tenhamos uma máquina do tempo para voltarmos e acompanharmos o passo a passo da evolução de determinada linhagem ao longo dos milhares de anos. Muitas críticas contra o uso dos fósseis foram levantadas na época. O próprio Hennig reconhecia que os dados fósseis poderiam ser úteis na hora de conduzir uma análise filogenética. Contudo, devido à tamanha incompletude do registro fossilífero, ou seja, tanta informação biológica era perdida no processo de formação de um fóssil, que estes deveriam ser preferivelmente menos utilizados na hora de reconstruir as relações de parentesco.
Petterson (1981) mostrou, através de vários exemplos, o quanto os fósseis prejudicavam a compreensão sobre as hipóteses de evolução dos animais. Ax em 1987 defendeu em seu livro The Phylogenetic System que os dados fósseis são tão incompletos que as árvores deveriam ser construídas com base apenas nos grupos viventes e só depois que a análise computacional fosse feita é que se deveriam adicionar os fósseis. Dessa maneira e com muito sucesso, os críticos rapidamente conseguiram marginalizar o uso dos dados paleontológicos nos estudos da evolução das linhagens de organismos viventes.
A Hipótese Clássica da evolução dos Amniotas
Inúmeras árvores foram construídas ao longo dos anos seguintes, a maioria delas ignorando as informações provindas dos fósseis. O estudo que mais me chamou a atenção foi o trabalho clássico de Gardiner em 1982, no qual este autor tentou reconstruir a até então, pouco compreendida história evolutiva dos amniotas. Amniota é um grupo de animais vertebrados que possuem, dentre muitas características, uma membrana extraembrionária ao redor do feto chamada de amnion, membrana a qual é fundamental para a independência da água do ambiente durante o desenvolvimento do filhote no ovo, permitindo que estes animais colonizassem completamente o habitat terrestre. Estamos falando então da maioria esmagadora de vertebrados terrestres (e os que secundariamente retornaram ao ambiente aquático) que dominaram a Terra: desde as formas extintas famosas como dinossauros, pterossauros, ictiossauros, plesiossauros até as espécies contemporâneas de tartarugas, crocodilos, lagartos, serpentes, aves e mamíferos.
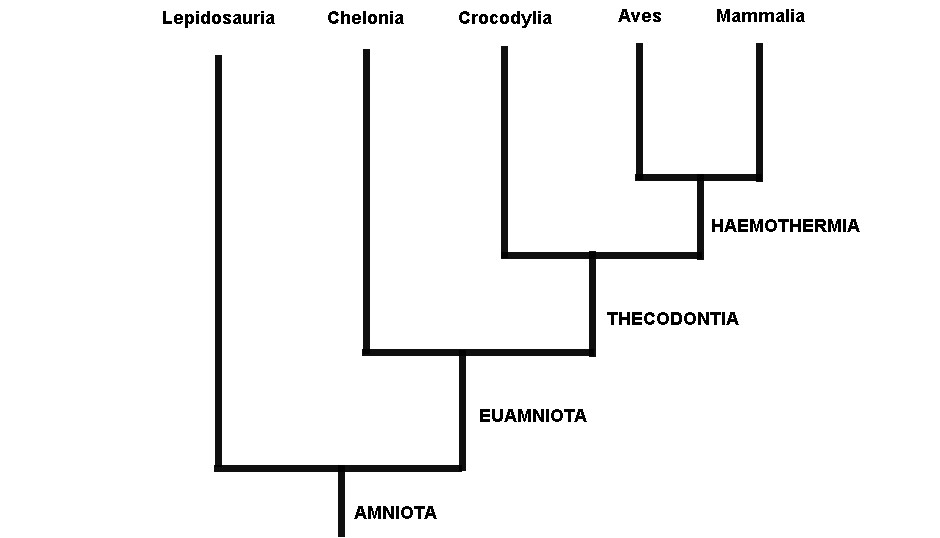
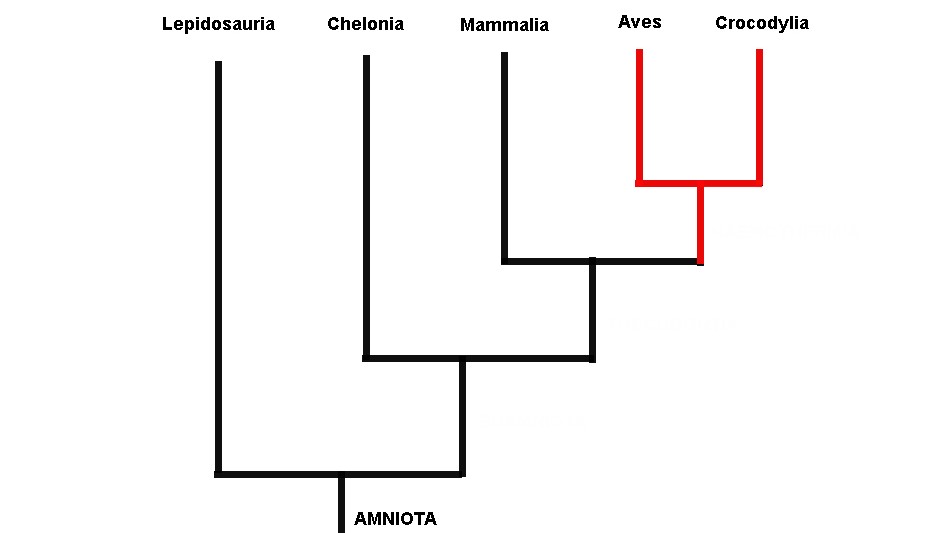
Gardiner utilizou vários dados morfológicos dos cinco grupos de amniotas viventes em suas análises (tartarugas, lagartos, jacarés, mamíferos e aves). Gardiner observou que as aves e os mamíferos eram evolutivamente relacionados, sendo agrupados por uma série de características que eram adaptações às suas altas taxas metabólicas (metabolismo alto leva a uma temperatura corpórea alta, sendo estes animais equivocadamente chamados de animais de “sangue quente”). Gardiner ressuscitou o antigo termo Haeomothermia para nomear o grupo de animais de “sangue quente” formado por aves e mamíferos (o termo vem de homeotermia, do grego homo: igual, thermia: temperatura; que é como chamamos os animais que possuem temperaturas corporais constantes). Haemothermia, por sua vez era relacionado evolutivamente com o Crocodylia (crocodilos, jacarés e gaviais) formando o grupo Thecodontia. Tartarugas, cágados e jabutis (Chelonia) eram mais aparentados com Thecodontia (Crocodylia+(Aves+Mammalia)). Por fim, o grupo mais basal de Amniota era Lepidosauria, o qual abrange tuataras, serpentes e lagartos (Figura 1).
Fig. 1. Relações entre as cinco assembleias de amniotas viventes defendida por Gardiner (1982; ver também Lovtrup, 1985) modificado de Gauthier e colaboradores (1988)
O trabalho experimental de Gauthier
Os resultados de Gardiner se baseam exclusivamente em animais viventes, conforme já foi dito e ecoaram por quase uma década. Lovtrup (1985) publicou um trabalho no qual afirmou ter encontrado maior suporte à hipótese de Gardiner. Até que, em 1988, um célebre manuscrito chegou para revolucionar a visão que a comunidade científica da época tinha sobre os dados paleontológicos. Gauthier e seus colegas (1988), baseados na hipótese de filogenia dos amniotas proposto por Gardiner, publicaram o primeiro estudo demonstrando empiricamente (ou seja, através de experimentos práticos e não calcados apenas em conjecturas) que os fósseis poderiam contribuir e muito na elucidação das hipóteses de parentesco dos seres vivos atuais. Para isso, eles conduziram uma nova análise, só que dessa vez incluindo espécies extintas, obtendo uma árvore marcadamente diferente e depois a submeteram a alguns testes para confirmar sua validade. Para entendermos como os autores chegaram a suas conclusões precisamos compreender os experimentos que a equipe realizou. Gauthier partiu de uma pergunta: os Fósseis poderiam alterar as nossas hipóteses de parentesco entre as biotas recentes? Para responder isso, ele conduziu um estudo em três etapas.
(1) Primeiramente, antes de rodar uma nova análise incluindo os fósseis, os pesquisadores destrincharam e reviram todas as características utilizadas por Gardiner e perceberam que havia alguns equívocos e erros na interpretação dos caracteres. Gardiner afirmara, por exemplo, que um coração dividido em quatro câmaras é homólogo (mesma origem; para melhor entendimento desta terminologia veja aqui) em crocodilos, aves e mamíferos, o que reforçava a relação de parentesco entre eles. Contudo, quando acompanhamos o desenvolvimento embrionário do septo interventricular destes animais, vemos que nos mamíferos ele se desenvolve a partir de uma crista de tecido endocárdico na parede de trás do ventrículo, enquanto que nas aves e nos crocodilos o septo surge de varias protuberâncias musculares pouco recobertas por endocárdio na lateral do ventrículo. Apesar de serem estruturas semelhantes nos adultos, elas não possuem a mesma origem no embrião, ou seja, não são homólogas. Logo, os equívocos nas interpretações de Gardiner o levaram a estabelecer homologias entre mamíferos e aves que não condiziam com a realidade.
(2) Depois de corrigir a lista de características, Gauthier rodou duas análises, uma contendo apenas os dados morfológicos dos cinco grupos viventes de Gardiner e outra incorporando 29 espécies de amniotas extintos. Foi então que uma nova hipótese começou a tomar forma.
A análise com dados dos animais viventes
A árvore filogenética obtida utilizando apenas os cinco grupos de amniotas viventes se assemelhou à hipótese clássica de Gardiner de 1982, com apenas uma pequena diferença: os crocodilos e não os mamíferos eram mais relacionados com as aves, trazendo dúvidas quanto a validade do antigo grupo Haemothermia (Ver figura 2). Isso implica em inferências importantíssimas para nossa compreensão da evolução da homeotermia. Primeiro, que um jacaré é o parente mais próximo das aves do que qualquer outro animal vivo hoje. Além disso, muitas características que antes eram vistas como homólogas entre aves e mamíferos e que estariam presentes no suposto ancestral comum destas duas linhagens, na verdade surgiram duas vezes independentemente na árvore da vida dos vertebrados. Talvez em resposta provavelmente as mesmas pressões evolutivas.
Fig. 2. Árvore obtida da análise de Gauthier (1988) apenas com amniotas viventes. Note que ela se assemelha à árvore anterior de Gardiner (1982), contudo as aves estão mais relacionadas com crocodilos do que com os mamíferos.
A análise com dados combinados (animais viventes + extintos)
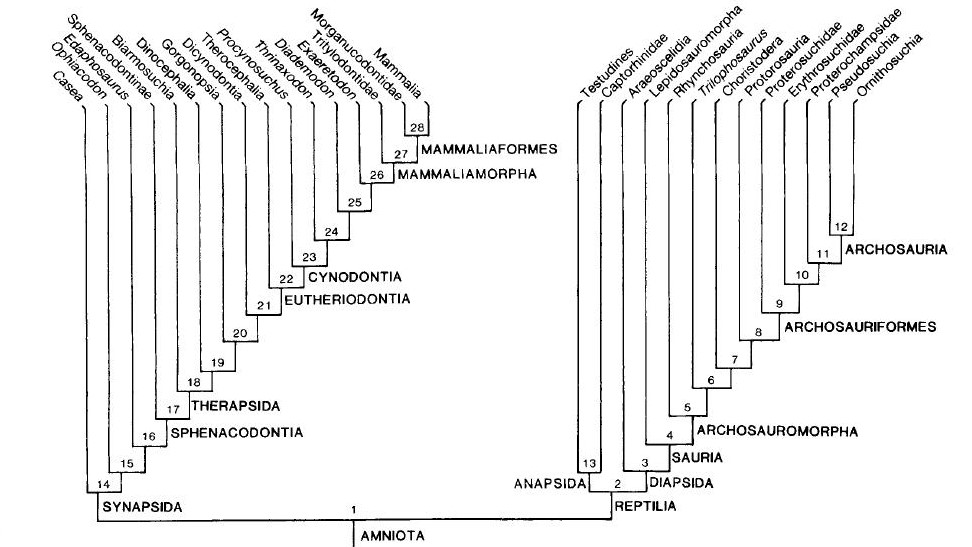
Na segunda análise na qual Gauthier acrescentou os fósseis, uma árvore completamente diferente surgiu (ver figura 3). Não só aves e crocodilos estavam agrupados juntos, mas lagartos e serpentes (Lepidosauria) agora estavam mais relacionados a Aves + Crocodylia do que as tartarugas. O mais estranho foi que os mamíferos agora estavam na base da árvore, formando uma grande dicotomia inicial: Mamíferos e todos seus parentes extintos de um lado versus lepidossauros, quêlonios, crocodilos, aves e todos seus parentes extintos relacionados do outro. Ao primeiro grupo, chamamos de Synapsida (amniotas que apresentam uma fenestra temporal no crânio) e ao segundo, Reptillia (que compreende tanto amniotas que possuem duas ou nenhuma fenestra temporal, Diapisida e Anapsida, respectivamente). Contudo, não bastava obter uma árvore completamente diferente, ela precisava ser mais bem justificada.
Fig. 3. Árvore obtidida por Gauthier (1988) combinando tanto espécies fósseis quanto viventes. Note que há uma dicotomia basal, separando mamíferos e seus parentes extintos (Synapsida) de um lado versus todos os répteis atuais + Aves (Reptilia) do outro.
Análises posteriores – Esmiuçando a nova hipótese obtida
Na última etapa do estudo (3), uma série de experimentos computacionais foram conduzidos por Gauthier, alguns serão explicados mais adiante e que culminaram em um achado esperançoso, principalmente para nós paleontólogos: Fósseis são fundamentais para elaboração das hipóteses de relação entre as espécies viventes. Gauthier e seus colegas queriam saber o porquê que as árvores diferiram tanto nas análises com e sem os fósseis. Para isso, eles fizeram algumas análises posteriores, contudo só irei detalhar três delas neste texto que acredito serem mais interessantes para nossa discussão. Primeiro, os autores compararam o índice de consistência de sua nova árvore com o índice da árvore de Gardiner. Este índice mostra o quão robusto e conciso estão seus resultados, no caso, sua hipótese de evolução de uma linhagem. O interessante foi que mesmo adicionando várias espécies fósseis na sua análise, Gauthier encontrou um valor de índice de consistência semelhante ao de Gardiner, mostrando que mesmo os fósseis alterando drasticamente a hipótese, a árvore continuava tão confiável quanto uma utilizando apenas espécies viventes.
Numa análise posterior, Gauthier e colaboradores removeram todos os grupos fósseis do lado “sinapsídeo” da árvore, deixando apenas os animais viventes desse ramo (mamíferos) junto com todos do lado “Reptillia” (viventes e extintos) e rodaram a análise. Paralelamente, foi feito o oposto, todas as linhagens extintas de Reptillia foram retiradas deixando apenas seus representantes viventes e o lado “sinapsídeo” da árvore (viventes e extintos) e rodaram a análise. Os autores viram que não importava qual fóssil de Reptillia fosse retirado, a árvore final não se alterava. Porém, quando os fósseis de sinapsídeos eram retirados, a árvore adquiria o novo padrão proposto por Gardiner (1982; Figura 3).
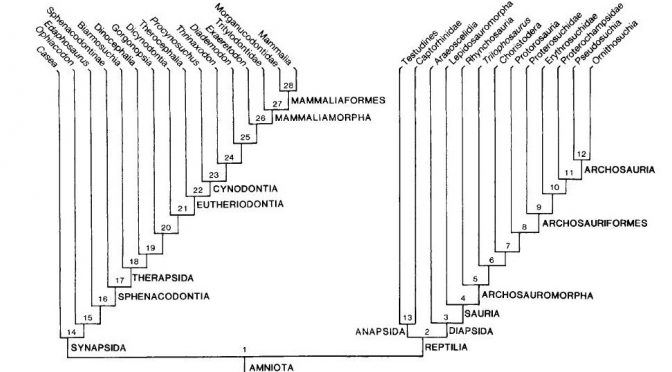
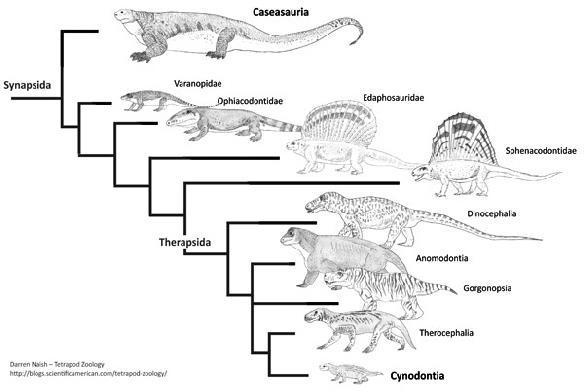
Gauthier então percebeu que os principais responsáveis pela nova topologia da árvore eram os fósseis de sinapsídeos. Mas por que isso? De acordo com os autores, quanto mais antiga é a origem de uma linhagem e quanto mais derivada for a morfologia de seus representantes atuais (como é o caso dos mamíferos), mais os fósseis serão importantes para elucidar sua evolução. Basta compararmos os mamíferos atuais com seus parentes extintos, os “pelicossauros” como Casea, Ophiacodon, Edaphosaurus, Sphenacodon (figura 4) para ver o “abismo morfológico” que separam essas linhagens. Quando observamos os mamíferos atuais, dificilmente conseguimos relaciona-los com outro grupo de animal vivente e quando tentamos, certamente estaremos fadados ao erro, assim como Gardiner, em 1982 equivocadamente agrupou Aves e Mammalia no antigo grupo Haeomothermia. Porém, quando olhamos para os fósseis vislumbramos um mundo completamente “novo”. As formas extintas possuem combinações únicas de características basais e derivadas que frequentemente se apresentam numa serie gradual de mudanças, que nos permitem acompanhar suas histórias evolutivas de maneira tal que possamos observar espécies diferentes ficando cada vez mais semelhantes conforme voltamos no tempo. Os fósseis são uma das poucas evidências diretas da evolução e é por isso, que os dados paleontológicos se tornam tão fundamentais para elucidar as relações evolutivas dos grupos viventes. Principalmente, quando o objeto de estudo são animais tão diferentes como seu cachorro e um pardal na janela de casa.
Outro experimento que a equipe de Gauthier conduziu consistiu basicamente em retirar todas as linhagens viventes da matriz de características e fazer uma nova análise. Apesar de ser um experimento simples, o resultado obtido foi revelador: a nova hipótese representada na figura 3 veio à tona novamente, porém sem os animais viventes. Este resultado, associado à análise inicial das características utilizadas no trabalho de Gardiner (1982), derruba os argumentos dos críticos a respeito da incompletude do registro fóssil. De fato a informação que provem dos fosseis é mais incompleta do que as que podemos retirar dos animais viventes, mas mesmo assim os fósseis estão longe de serem menos informativos em uma análise filogenética. Gauthier demonstrou isso quando retirou todas as espécies viventes da análise e mesmo assim alcançou a nova hipótese. Além disso, Gauthier enfatizou que incompletude não é exclusiva de fósseis. Ela pode ocorrer naturalmente nos animais. Quatro das características utilizadas na análise de Gardiner eram do osso quadrado, um osso na base do crânio dos vertebrados no qual se articula a mandíbula. Contudo, nos sinapsídeos mais derivados, os cinodontes (o qual mamíferos fazem parte), esse osso foi gradativamente sendo reduzido e alocado cada vez mais para trás do crânio junto com uma série de ossos da mandíbula. Esta condição alcançou o extremo nos mamíferos, onde o osso quadrado e um grupo de ossos que uma vez pertenceram à mandíbula, se reduziram e modificaram sua função ao ponto de formarem o que hoje são nossos ossículos do ouvido médio (o quadrado dos outros vertebrados é a bigorna nos mamíferos). Logo, aquelas quatro características baseadas na morfologia do osso quadrado, não se aplicam aos mamíferos, pois o quadrado deles se alterou tanto que fica difícil comparar com os outros animais, ou seja, este é um dado naturalmente faltante. Além disso, Gauthier percebeu que Casea (o fóssil mais basal e antigo de sinapsídeo e que consequentemente esperávamos maior incompletude) apresentava 26% de informação faltando, enquanto que os mamíferos atuais tinham em média 15% de dados morfológicos faltando ou difíceis de interpretar. Esses 11% de diferenças são realmente significantes? Gauthier não só provou que não há diferença significante como também nos mostrou que informação incompleta não é exclusiva dos fósseis.
Fig. 4. Outra árvore filogenética, dessa vez apenas com sinapsídeos. Note a variedade de formas desde as mais basais, como Casesauria (um “Pelicossauro”), até os parentes mais próximos dos mamíferos, um cinodonte não-mamífero.
A Renascença dos fósseis
Após demonstrar empiricamente quão importante os fósseis são para nossa compreensão da evolução das linhagens viventes, Gauthier deu um solavanco nos sistematas, convidando-os a se debruçarem no assunto. Desde então, estudos importantes vêm sendo conduzidos, cada vez mais enaltecendo o uso de dados paleontológicos em reconstruções filogenéticas e desenvolvendo metodologias para minimizar o efeito da incompletude dos dados de organismos tanto viventes quanto extintos, o famigerado missing data que os cladistas tanto abominam (veja Donoghue et al., 1989; Smith, 1998; Wilkinson; Benton, 1995). Exemplos como estes de Gardiner e Gauthier nos mostram como a Ciência é dinâmica. O que antes era tido como verdade e que hoje é obsoleto, não necessariamente deixa de ser útil. Se Gardiner não tivesse se aventurado nas relações de Amniota, Gauthier e toda uma geração de sistematas não seriam impulsionadas a refletir sobre o assunto e talvez o reconhecimento da importância dos fósseis na reconstrução da evolução da vida na Terra poderia ser ainda mais postergada. Por fim, quando ignoramos as informações contidas no registro fóssil, estamos ferindo o princípio da Evidência Total (um tema que será abordado em postagens futuras do blog). Este princípio pode ser exemplificado com uma ótima analogia que um estimado amigo uma vez me fez: não usar os dados paleontológicos para inferir filogenia é como ter um bebê de colo e joga-lo pela janela só porque ele não é um adulto.
Referências:
AX, Peter. 1987. The phylogenetic system: the systematization of organisms on the basis of their phylogenesis. DARWIN, C. A Origem das Espécies. Hemus – Livraria Editora Ltda, São Paulo, SP.
DONOGHUE, Michael J. 1989. Phylogenies and the analysis of evolutionary sequences, with examples from seed plants. Evolution, v. 43, n. 6, p. 1137-1156.
GARDINER, BRIAN G. 1982. Tetrapod classification. Zoological Journal of the Linnean Society, v. 74, n. 3, p. 207-232.
GAUTHIER, Jacques; KLUGE, Arnold G.; ROWE, Timothy. 1988. Amniote phylogeny and the importance of fossils. Cladistics, v. 4, n. 2, p. 105-209. HENNIG, Willi. 1950. Grundzuge einer Theorie der phylogenetischen Systematik.
LOVTRUP, Soren. 1985. On the classification of the taxon Tetrapoda. Systematic Zoology, v. 34, n. 4, p. 463-470.
PATTERSON, Colin. 1981. Significance of fossils in determining evolutionary relationships. Annual Review of Ecology and Systematics, v. 12, n. 1, p. 195-223.
SMITH, Andrew B. 1998. What does palaeontology contribute to systematics in a molecular world?. Molecular phylogenetics and evolution, v. 9, n. 3, p. 437-447.
WILKINSON, Mark; BENTON, Michael J. 1995. Missing data and rhynchosaur phylogeny. Historical Biology, v. 10, n. 2, p. 137-150.
Geovane Alves de Souza, Graduado em licenciatura e bacharelado em Ciências Biológicas pela Universidade Estadual de Londrina. Atualmente é mestrando em Zoologia pelo Museu Nacional/UFRJ. Já desenvolveu pesquisas na área de parasitologia de animais silvestres, hoje conduz estudos de osteohistologia com titanossauros.
Novo estudo, liderado por pesquisador brasileiro e publicado hoje na revista científica ‘Scientific Reports‘ (UK), investigou o cérebro de um dos mais antigos dinossauros do mundo e revelou detalhes importantes sobre o comportamento desses animais. O dinossauro estudado foi Saturnalia tupiniquim, uma espécie brasileira, descoberta no Rio Grande do Sul, e proveniente de unidade geológica de relevância internacional, que tem fornecido peças chaves na compreensão da evolução dos primeiros dinossauros.
Reconstituição de Saturnalia tupiniquim, arte de Rodolfo Nogueira.
O estudo foi liderado pelo aluno de doutorado Mario Bronzati Filho, do Programa Ciência sem Fronteiras (CNPq), e contou ainda com a participação do pesquisador alemão Dr. Oliver W. M. Rauhut (Ludwig-Maximilians-Universität), supervisor do estudante na Alemanha, e também com professores de duas universidades brasileiras, Dr. Jonathas S. Bittencourt (UFMG) e Dr. Max C. Langer (FFCLRP-USP).
Os pesquisadores usaram microtomografia computadorizada para reconstruir o cérebro de Saturnalia e, assim, puderam entender em maior detalhe a sua estrutura. Com base nisso, foi possível realizar inferências sobre o comportamento desses animais. Esta foi a primeira vez que partes do cérebro de um dinossauro tão antigo foram reconstruídas. Os fósseis de Saturnalia foram encontrados em rochas de 230 milhões de anos da chamada Formação Caturrita, unidade geológica da Bacia Sedimentar do Paraná, que afloram no estado do Rio Grande do Sul.
Afloramento rochoso da Formação Caturrita onde foram encontrados os fósseis de Saturnalia Tupiniquim.
Saturnalia tupiniquim foi um dinossauro de pequeno porte, com cerca de 1,5 m de comprimento e que pesava aproximadamente 10 kg. Seus fósseisforam encontrados há cerca de 20 anos, na área urbana de Santa Maria, RS. Muitos detalhes da anatomia do seu esqueleto pós-craniano (isto é, dos membros, coluna vertebral e cinturas escapular e pélvica)já eram conhecidos, mas essa foi a primeira vez que partes internas de seu crânio foram estudadas.
Reconstituição artística de Saturnalia tupiniquim, arte de Rodolfo Nogueira (http://rodolfonogueira.daportfolio.com).
Reconstrução esqueletal e escala de tamanho de Saturnalia tupiniquim. Arte de Felipe A. Elias.
Saturnalia pertence ao grupo de dinossauros conhecido como ‘Sauropodomorpha’, uma linhagem de espécies essencialmente herbívoras, que inclui os famosos saurópodes, dinos de pescoço comprido, que estão entre alguns dos maiores animais que já caminharam sobre o planeta Terra, como os titanossauros, o apatossauro e o braquiossauro. A linhagem Sauropodomorpha viveu na Terra por cerca de 170 milhões de anos e os últimos representantes desse grupo foram extintos há cerca de 66 milhões, durante o evento de extinção em massa do final do Cretáceo, que extinguiu todos os dinossauros não-avianos (as Aves, que são dinossauros, continuam muito bem, obrigada).
Sauropodomorpha, arte de Franz Anthony.
Saturnalia é um dos mais antigos Sauropodomorpha conhecidos (no que diz respeito à tamanho, essa foi uma origem bastante humilde para um grupo de gigantes, não?). O resultado curioso do estudo de Bronzati e colegas é que, com base em análises comparativas, o cérebro de Saturnalia revela que ele – literalmente – tinha “a cabeça de um predador”. Isso é extremamente relevante do ponto de vista ecológico e evolutivo. A linhagem dos Sauropodomopha é conhecida pela evolução de formas muito bem adaptadas à herbivoria, porém o estudo com Saturnalia revela que a origem desse grupo de dinossauros estava longe de ser “vegana”…
A dieta de um dinossauro é geralmente inferida com base na morfologia dos dentes e de outras partes de seu esqueleto. Entretanto, a morfologia do cérebro também pode fornecer informações valiosas para o estudo do comportamento de animais extintos. Diferentemente dos gigantes saurópodes, que tinham uma dieta baseada somente em plantas, a dentição dos primeiros sauropodomorfos (incluindo Saturnalia tupiniquim) indica que esses animais tinham uma dieta onívora (baseada em plantas e outros animais) ou faunívora (baseada somente em outros animais). Agora, a nova pesquisa traz evidências adicionais deque os sauropodomorfos mais antigos eram animais predadores.
A observação exclusiva dos dentes de Saturnalia não era muito conclusiva sobre a sua dieta. O novo estudo traz informações melhor embasadas. Frame do vídeo de divulgação do artigo, arte de Rodolfo Nogueira.
De acordo com a avaliação dos paleontólogos responsáveis pelo estudo, Saturnalia apresentava um grande volume do “flóculo e o paraflóculo do cerebelo”. Esqueça os nomes complicados… Estes tecidos fazem parte de sistemas neurológicos que operam no controle do movimento de cabeça e do pescoço do animal, e também no controle da visão. O grande volume destas estruturas indica um comportamento em que movimentos rápidos de pescoço e cabeça deveriam ser parte do repertório desse animal. Isso é observado, normalmente, em predadores, que usam essas habilidades para capturar presas pequenas e esquivas. Apesar do alongamento do pescoço e a redução do crânio representarem marcas registradas do plano corpóreo Sauropodomorpha mais derivados (i.e. os saurópodes), os primeiros passos na aquisição dessa morfologia única parecem ter surgido como adaptações para predação, um cenário evolutivo conhecido como exaptação, explicam os autores.
Exaptação é um processo em que uma característica surge com uma certa função, mas passa a ter outra função em um momento distinto da história evolutiva de uma linhagem.
Os saurópodes, herbívoros, não possuíam grandes volumes dos tecidos do cérebro ligados ao controle refinado do movimento da cabeça, pescoço e visão. Isto indica que a evolução da herbivoria nessa linhagem também se deu com mudanças na estrutura dos cérebros desses animais.
O estudo do comportamento de animais extintos é muito difícil e muitas vezes depende da observação de evidências indiretas, como os icnofósseis. Estudos de reconstruções digitais do cérebro são uma forma de se obter tal tipo de informação, mas eles sempre devem ser interpretados com cautela. Não obstante, o novo estudo é um importante primeiro passo na busca por uma melhor compreensão do comportamento dos primeiros dinossauros. Por fim, estudos futuros certamente trarão mais informações para entender em mais detalhes a evolução da linhagem dos sauropodomorfos, que começou com pequenos animais predadores e posteriormente deu origem aos gigantes herbívoros do passado.
Reconstituição em vida de Saturnalia tupiniquim. Arte de Rodolfo Nogueira.
Veja o vídeo de divulgação do artigo:
Veja algumas informações exclusivas no bate-papo que tivemos com o Mario Bronzati, autor principal do estudo, em nosso canal (clique na imagem para ser redirecionado para o YouTube):
Agradecemos ao colega Mário Bronzati por ter compartilhado as informações desse maravilhoso estudo conosco antecipadamente. Ademais, não deixem de se inscrever em nosso canal (http://www.youtube.com.br/colecionadoresdeossos)!
Bronzati-Filho et al. 2017. Endocast of the Late Triassic (Carnian) dinosaur Saturnalia tupiniquim: implications for the evolution of brain tissue in Sauropodomorpha. Scientific Reports.