


Beatas no Ninho
Embora o título possa parecer um pouco reles, o teor deste texto é tudo menos provocador.
Bem, talvez o seja para alguns seres vivos.
As beatas de que falamos são as pontas dos cigarros depois de fumados ou, em português do Brasil, as bitucas ou guimbas de cigarro.
 Mas porquê falar da terminologia de um produto tão nocivo à saúde e, pior, misturá-lo com ninhos?
Mas porquê falar da terminologia de um produto tão nocivo à saúde e, pior, misturá-lo com ninhos?
É que por vezes a natureza dá voltas por onde menos se espera. Neste caso, verificou-se que algumas aves da cidade do México utilizam as beatas de cigarros na construção dos seus ninhos.
O comportamento foi observado e avaliado por investigadores da Universidad Nacional Autónoma de México, que utilizaram um procedimento experimental para comprovarem a influência das guimbas de cigarro sobre parasitas que atacam as crias de aves.
Os cientistas verificaram que ninhos com pontas de cigarros fumados apresentavam menos parasitas externos do que aqueles construídos com pontas de cigarros não fumados.
Bem, nada de especial, escarnecerão os mais radicais, acrescentando que o tabaco é tão mau que nem os parasitas o aguentam.
 Os investigadores verificaram que as aves que utilizam as beatas nos seus ninhos, pardais (Passer domesticus) e a espécie de tentilhão Carpodacus mexicanus, o poderão fazer como recurso a um insecticida natural, já que as pontas dos cigarros preservam quantidades de nicotina e outras substâncias químicas.
Os investigadores verificaram que as aves que utilizam as beatas nos seus ninhos, pardais (Passer domesticus) e a espécie de tentilhão Carpodacus mexicanus, o poderão fazer como recurso a um insecticida natural, já que as pontas dos cigarros preservam quantidades de nicotina e outras substâncias químicas.
Um amigo meu inglês já me havia descrito que infusões frias de beatas de cigarros quando vertidas nos vasos de plantas ornamentais as tornam mais saudáveis. Ou assim diz ele.
Estes investigadores não descartaram a hipótese de que as aves utilizem as beatas como um revestimento térmico para os ninhos, uma vez que estas têm celulose. Tão pouco afastam que as vantagens de as aves utilizarem a nicotina como desparasitante sejam anuladas pelos efeitos tóxicos dos químicos tabágicos.
 Ainda que não totalmente esclarecidos, os autores propõem duas hipóteses para o comportamento das aves: as substâncias químicas presentes nas pontas de cigarro poderão estimular o sistema imunitário das crias e, assim, favorecer as suas hipóteses de sobrevivência. A segunda hipótese aponta para que os químicos presentes nas bitucas de cigarros possam ter um papel mais directo, evidente e já referido: as beatas seriam um insecticida natural, que desinfectaria os ninhos de parasitas externos.
Ainda que não totalmente esclarecidos, os autores propõem duas hipóteses para o comportamento das aves: as substâncias químicas presentes nas pontas de cigarro poderão estimular o sistema imunitário das crias e, assim, favorecer as suas hipóteses de sobrevivência. A segunda hipótese aponta para que os químicos presentes nas bitucas de cigarros possam ter um papel mais directo, evidente e já referido: as beatas seriam um insecticida natural, que desinfectaria os ninhos de parasitas externos.
Esquecendo as infusões de nicotina e desejando que as hipóteses levantadas sejam testadas, o que as aves urbanas parecem ter descoberto é a reutilização do arsenal químico dos cigarros a favor das suas crias.
Ao contrário do que escreveu Tchekov, estas aves mexicanas descobriram os benefícios do tabaco.
Aparentemente.
![]() Referências:
Referências:
Suárez-Rodríguez M, López-Rull I, & Macías Garcia C (2012). Incorporation of cigarette butts into nests reduces nest ectoparasite load in urban birds: new ingredients for an old recipe? Biology letters, 9 (1) PMID: 23221874
Imagens:
1 – daqui
2 – de Vitor Argaez – daqui
3 – daqui
Disparidade e Biodiversidade em Dinossauros
Este texto foi escrito na sequência do contacto do jornalista Marco Túlio Pires da revista brasileira Veja, que pretendia um comentário meu ao artigo da Nature Communications “Dinosaur morphological diversity and the end-Cretaceous extinction”. (ver Abstract abaixo)
Este foi o texto que enviei e que foi parcialmente citado no artigo publicado na revista Veja on-line.
 A questão fundamental que este artigo encerra é:
A questão fundamental que este artigo encerra é:
a) qual o estado ecológico de Dinosauria, ou seja, a sua biodiversidade, aquando da queda do meteorito há 65 milhões de anos, ou seja, estariam já as espécies de dinossauros do final do Cretácico em declínio no momento da queda do meteorito, tendo este apenas acelerado a sua extinção?
A questão não é nova, tendo sido analisada por muitos paleontólogos nas últimas décadas.
Este artigo é relevante pois analisa uma quantidade grande de grupos de dinossauros, aplicando uma série de metodologias de medição da disparidade morfológica que, para os autores deste estudo, é sinónimo de biodiversidade.
A metodologia utilizada por estes investigadores assenta na medição da variabilidade de formas anatómicas destes animais – disparidade morfológica, como indicador da biodiversidade de determinado grupo.
Neste perspectiva este estudo é inovador, deduzindo que alguns grupos de dinossauros estariam em melhor estado ecológico do que outros nos últimos 12 milhões de anos do Cretácico.
Assim, dinossauros carnívoros e herbívoros de porte pequeno a médio, como paquicefalossauros e anquilossauros, mantiveram a sua disparidade morfológica (biodiversidade) durante o final do Cretácico.
Pelo contrário, os autores do artigo verificaram que os grandes herbívoros como os hadrossaurídeos e ceratopsídeos estavam em declínio ao nível da sua disparidade morfológica nos últimos 12 milhões de anos do Mesozóico.
Há ainda o caso dos saurópodes, os maiores dinossauros, quadrúpedes e de cauda e pescoços compridos, que apresentam um aumento da sua biodiversidade antes mesmo da queda do meteorito. Os saurópodes pareciam estar destinados a um futuro brilhante não fosse a queda do meteorito…
 A minha tese de doutoramento (Universidad Autónoma de Madrid, 2009) quantificou a disparidade morfológica dos ossos das patas de Sauropodomorpha, e outros grupos como terópodes, aves, pterossauros e morcegos, revelou que o aumento de tamanho neste grupo coincidia com um aumento na disparidade morfológica das patas destes animais.
A minha tese de doutoramento (Universidad Autónoma de Madrid, 2009) quantificou a disparidade morfológica dos ossos das patas de Sauropodomorpha, e outros grupos como terópodes, aves, pterossauros e morcegos, revelou que o aumento de tamanho neste grupo coincidia com um aumento na disparidade morfológica das patas destes animais.
Por outras palavras, quanto maiores forem os saurópodes, maior a disparidade morfológica do esqueleto apendicular destes animais.
Tenho utilizado a quantificação da disparidade morfológica, no caso dos ossos das patas, mas com objectivos distintos deste estudo. O meu interesse é perceber, por exemplo, porque é que e como é que alguns dinossauros mudaram o seu modo de locomoção de bípedes para quadrúpedes.
Apesar de importante, o artigo de Brusatte et al. não responde a algumas questões fundamentais, tais como:
– estavam os dinossauros em declínio no momento da queda do meteorito? Uns estavam, outros não, e outros nem por isso.
-qual o motivo da extinção diferencial no final do Cretácico, ou seja, porque o impacto do meteorito originou a extinção de umas espécies (por exemplo dinossauros e outros animais terrestres, mas também amonites, belemnites e outra fauna aquática) e não originou a extinção de outras (como mamíferos, aves, entre outras)?
– quais os resultados deste tipo de análise com aumento da amostra e com fauna de outros locais? Manter-se-iam os resultados? Os dados analisados são sobretudo, mas não exclusivamente, de formas norte-americanas embora também existam de outras áreas geográficas.
Em resumo: este artigo é importante porque quantifica a biodiversidade dos dinossauros do final do Cretácico, embora o seja mais por apontar um caminho para o futuro da investigação paleobiológica em dinossauros que é o da quantificação da disparidade morfológica como indicador da biodiversidade (neste caso a paleodiversidade).
A Extinção do final do Cretácico
Neste momento a teoria mais aceite para explicar a extinção dos dinossauros não-avianos (e de muitas outras espécies) é a queda de um objecto extra-terrestre.
Apesar de não conseguir explicar a extinção diferencial que se verificou no final do Cretácico, ou seja, por que é que umas espécies se extinguiram e outras não, a teoria do extinção por impacto, apresentada na década de 80 do século passado, está muito bem comprovada.
Como?
Sabe-se que o metal irídio é um metal de muito baixas concentrações na Terra. O que Walter Alvarez e colegas detetaram foi concentrações muito superiores ao normal em sedimentos do final do Cretácico.
O que significa isto?
Que um objeto, como um meteoro, que apresentam concentrações de irídio muito superiores às da Terra deveria ter chocado com o nosso planeta.
As provas deste choque, ou seja, as concentrações anómalas de irídio foram encontradas em vários pontos da Terra permitindo comprovar aquela impacto.
Outras provas da queda, como a cratera de impacto, demoraram mais tempo a serem encontradas. Um dos possíveis locais de queda e que sempre foi apontado foi a península do Iucatão, no México (Chicxulub). O que é certo é que 65 milhões de anos de erosão alteraram a forma inicial da cratera, tornando mais difícil a sua identificação.
Recentemente, alguns autores responsabilizaram a queda do asteróide Baptistina como a origem da mega-extinção que ocorreu há 65 milhões de anos, no final do Cretácico. Este ano, estudos da NASA descartaram esta hipótese uma vez que parte da fragmentação de Baptistina atingiu a Terra mais cedo, ou seja, há 80 milhões de anos.
Entre a comunidade científica existem outras teorias para justificar a extinção, embora menos aceites que a da queda de um objecto extra-terrestre. Alguns cientistas, apontam a ocorrência de actividade vulcânica de grande intensidade e que ao libertarem milhões de toneladas de gases poderão ter contribuído para o desaparecimento em massa de seres vivos. Esta hipótese não exclui a da queda de um meteorito e, provavelmente, a explicação da extinção do final do Cretácico será terá apenas uma causa mas a combinação de vários motivos – queda de meteorito, vulcanismo e outras.
Abstract do artigo de Brusatte et al.
“The extinction of non-avian dinosaurs 65 million years ago is a perpetual topic of fascination, and lasting debate has focused on whether dinosaur biodiversity was in decline before end-Cretaceous volcanism and bolide impact. Here we calculate the morphological disparity (anatomical variability) exhibited by seven major dinosaur subgroups during the latest Cretaceous, at both global and regional scales. Our results demonstrate both geographic and clade-specific heterogeneity. Large-bodied bulk-feeding herbivores (ceratopsids and hadrosauroids) and some North American taxa declined in disparity during the final two stages of the Cretaceous, whereas carnivorous dinosaurs, mid-sized herbivores, and some Asian taxa did not. Late Cretaceous dinosaur evolution, therefore, was complex: there was no universal biodiversity trend and the intensively studied North American record may reveal primarily local patterns. At least some dinosaur groups, however, did endure long-term declines in morphological variability before their extinction.”
Referências:
Stephen L. Brusatte, Richard J. Butler, Albert Prieto-Márquez & Mark A. Norell. 2012. Dinosaur morphological diversity and the end-Cretaceous extinction. Nature Communications 3, Article number: 80 doi:10.1038/ncomms1815.
Rodrigues, L.A. Sauropodomorpha (Dinosauria, Saurischia) appendicular skeleton disparity: theoretical morphology and Compositional Data Analysis. Universidad Autónoma de Madrid, Madrid – Spain, Supervised by Professor Angela Delgado Buscalioni and Co-supervised by Professor Jeffrey A. Wilson, University of Michigan, Ann Arbor. December 2009. ISBN 978-84-693-3839-1.
Imagens:
De Brusatte et al 2012 e de Rodrigues, L.A. 2009
Tento na Língua
 Nenhum dos casos é muito agradável, muito menos para mentes eticamente bem formadas ou estômagos sensíveis.
Nenhum dos casos é muito agradável, muito menos para mentes eticamente bem formadas ou estômagos sensíveis.
Ainda assim, encontro-lhes pontos de contacto.
![]()
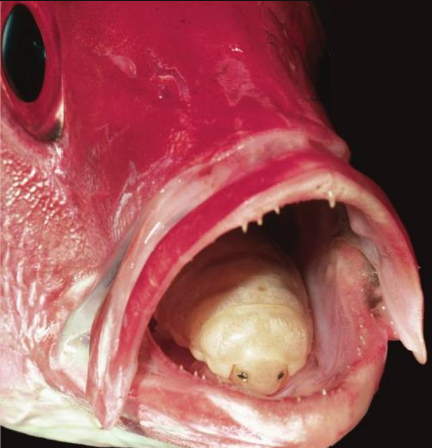
O parasita Cymothoa exigua é um crustáceo isópode que actua de maneira singular.
Explico, enquanto se torcem.
Já na boca do peixe, o C. exigua fixa-se na boca do peixe por intermédio de patas semelhantes a ganchos (pereópodes), começando por sugar o sangue dos tecidos da língua, até que esta acaba por atrofiar.
Após o repasto, que pode durar algum tempo, o parasita de até quatro centímetros, passa a alimentar-se do que o peixe ingere, substituindo-lhe a língua por completo.
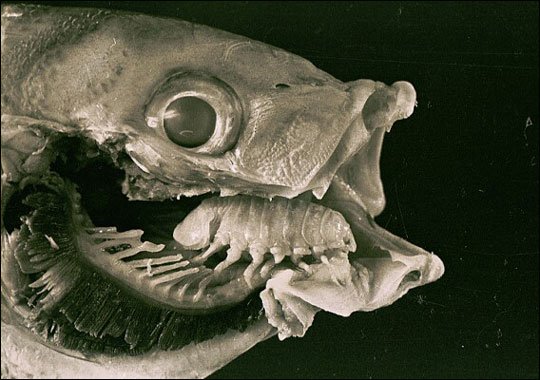 Curiosa é a semelhança de forma entre a desaparecida língua do peixe e o recém instalado. Para além de função análoga, já que o parasita desempenha a usurpação lingual com enorme competência, também o aspecto da cavidade bucal do peixe parece quase inalterada. Brusca e Gilligan avançam com a hipótese de que peixes parasitados poderão ter melhor desempenho alimentar que peixes sem língua.
Curiosa é a semelhança de forma entre a desaparecida língua do peixe e o recém instalado. Para além de função análoga, já que o parasita desempenha a usurpação lingual com enorme competência, também o aspecto da cavidade bucal do peixe parece quase inalterada. Brusca e Gilligan avançam com a hipótese de que peixes parasitados poderão ter melhor desempenho alimentar que peixes sem língua.
Ou seja, em termos de eficiência, esta relação parasítica parece conceder alguma vantagem sobre patologia ou doenças que afectem a língua de peixes.
Pois…melhor uma língua substituta que nenhuma, parece ser a conclusão.
Quanto ao segundo caso, apenas deixo um vídeo.
Deixo aos leitores o estabelecimento de paralelismos.
A existirem.
Referências:
Brusca, R.C. and Gilligan, M.R. (1983): Tongue replacement in a marine fish (Lutjanus guttatus) by a parasitic isopod (Crustacea: Isopoda). Copeia 813-816.
Brusca, R., & Gilligan, M. (1983). Tongue Replacement in a Marine Fish (Lutjanus guttatus) by a Parasitic Isopod (Crustacea: Isopoda) Copeia, 1983 (3) DOI: 10.2307/1444352
Mark Carwardine. 2005.Natureza radical. Ediouro Publicações Ltda. Rio de Janeiro.
Zimmer, C. 2000. Parasite Rex. Arrow Books. London.
Imagens:
Matthew Gilligan – primeira, adaptada do livro de Mark Carwardine; segunda daqui
Biologia Evolutiva
Entrevista ao Biólogo Evolutivo, André Levy, conduzida por alunos do 12º ano da Escola Secundária Henriques Nogueira, Torres Vedras.
Este grupo de alunos desenvolveu os seus trabalhos de Área de projecto no tema da Medicina Evolutiva.
Paleo talk ou Miragaia longicollum
 Logo após a publicação do artigo científico que descrevia um nada usual estegossáurio, enviei um pequeno questionário ao paleontólogo Octávio Mateus.
Logo após a publicação do artigo científico que descrevia um nada usual estegossáurio, enviei um pequeno questionário ao paleontólogo Octávio Mateus.
Pretendia que Mateus, numa linguagem acessível, descrevesse o Miragaia longicollum bem como a importância deste exemplar para a compreensão da vida passada na Terra.
Aqui está a mini-entrevista ou, como prefiro, uma troca de mails entre paleontólogos:
Ciência Ao Natural (CAN): Como apresentaria este novo dinossáurio ao público?
Octávio Mateus (OM): Como todos os dinossauros estegossauros é um quadrúpede herbívoro, com placas no dorso, espinhos na cauda e membros anteriores curtos. Mas ao contrário dos outros estegossauros, este tem um pescoço invulgarmente longo, com 17 vértebras cervicais, o que representa 10 a mais do que a girafa e o maior número entre todos os dinossauros não-avianos.
CAN: Esqueça que participou nesta descoberta. Como classificaria a importância deste fóssil?
OM: Este fóssil é um dos dinossauros portugueses mais completos e o facto de ser um novo taxon vem reforçar a importância de Portugal no cenário da riqueza mundial de dinossauros. A sua inesperada característica de pescoço longo leva-nos a uma série de questões fascinantes sobre a evolução destes dinossauros. Além disso, o Miragaia longicollum demonstra a plasticidade evolutiva dos dinossauros.
 CAN: As semelhanças morfológicas, ao nível do esqueleto do pescoço, entre dois grupos de dinossáurios distintos como os saurópodes e o estegossáurios são evidentes nunca antes registadas. Que podemos inferir destas parecenças, ao nível da evolução dos dinossáurios?
CAN: As semelhanças morfológicas, ao nível do esqueleto do pescoço, entre dois grupos de dinossáurios distintos como os saurópodes e o estegossáurios são evidentes nunca antes registadas. Que podemos inferir destas parecenças, ao nível da evolução dos dinossáurios?
OM: As semelhanças mostram que a convergência evolutiva é tão surpreendente como lógica e expectável. Ou seja, se por um lado ninguém esperava ver um estegossauro com 17 vértebras cervicais e superficialmente parecido a um saurópode, por outro lado, a plasticidade evolutiva associada à necessidade de adaptação ao mesmo ambiente dos saurópodes faz da convergência um processo comum e expectável.
CAN: Qual o dinossáurio que gostaria de descobrir em Portugal? Porquê?
OM: Gostaria de descobrir algo que não estivesse à espera de o fazer. Quanto mais inusitado, mais interessante será. Uma descoberta verdadeiramente inesperada levantaria muitas mais questões científicas e tornaria a paleontologia ainda mais cativante.
CAN: Qual o conselho que daria a um jovem português que gostasse de seguir as “pisadas” de Octávio Mateus?
OM: Sem me considerar um modelo, acho que o desejo de aprender mais sobre história natural é a minha grande motivação. Aconselho a interessarem-se pela história natural e nunca perderem a vontade de estudar.
Referência: Mateus, O., Maidment, S.C.R., and N.A. Christiansen. 2009. A new long-necked ‘sauropod-mimic’ stegosaur and the evolution of the plated dinosaurs. Proceedings of the Royal Society B, first online. DOI 10.1098/rspb.2008.1909.
Imagens: do blog Lusodinos
I Iberian Symposium on Geometric Morphometrics
 Há quase cinco anos organizei e co-ministrei um Workshop de Morfometria Geométrica – resumo aqui. Agora é a vez de divulgar o I Iberian Symposium on Geometric Morphometrics, que decorrerá na cidade catalã de Sabadell, entre 23 e 25 de Julho de 2009.
Há quase cinco anos organizei e co-ministrei um Workshop de Morfometria Geométrica – resumo aqui. Agora é a vez de divulgar o I Iberian Symposium on Geometric Morphometrics, que decorrerá na cidade catalã de Sabadell, entre 23 e 25 de Julho de 2009.
Agradeço o honroso convite da organização para fazer parte da Comissão Científica, especialmente porque esta integra investigadores de excelente nível, como F. James Rohlf (State University of New York), Christian Klingenberg (University of Manchester), Paul O’Higgins (Hull-York Medical School, University of York), Diego Rasskin-Gutman (Universidad de Valência), entre outros.
A Morfometria Geométrica é um conjunto de técnicas de análise de variação da forma orgânica.
 “The advent of Geometric Morphometrics was claimed as a revolution at the end of the century, and step by step it has grown to become a customary tool to analyze and understand the ultidimensional nature of biological form.
“The advent of Geometric Morphometrics was claimed as a revolution at the end of the century, and step by step it has grown to become a customary tool to analyze and understand the ultidimensional nature of biological form.
This symposium is organized to gather for the first time all practitioners of the field, as a way to share each one´s knowledge, findings and concerns, but also as a way to begin opening a window for future collaborations.”
(do site do Simpósio)
Um FAQ da Universidade de Viena sobre a Morfometria Geométrica.

Informações:
Imagens – dos sites do Simpósio e da Universidade de Viena.
II O canário do mineiro
 (continuação)
(continuação)
 São referidos números distintos mas é unânime que os números avançados, em 1993, por E.O. Wilson – que cerca de 30 000 espécies desaparecem por ano – se encontram desactualizados, mas por defeito…
São referidos números distintos mas é unânime que os números avançados, em 1993, por E.O. Wilson – que cerca de 30 000 espécies desaparecem por ano – se encontram desactualizados, mas por defeito… Porque devemos então preocupar-mo-nos que uma espécie de anfíbio da América do Sul se extinga?
Porque devemos então preocupar-mo-nos que uma espécie de anfíbio da América do Sul se extinga?Referências
existe vasta bibliografia referente a extinções em massa; recomendo sobretudo os trabalhos de Douglas Erwin, que há mais 15 anos teve a simpatia e modéstia de responder directamente a uma questão trivial de um estudante de licenciatura sobre …o estudante era eu; a questão tenho pudor em a revelar.
Imagens (dos dois posts e por ordem):
daqui, daqui, daqui, daqui, daqui e, finalmente daqui.
O canário do mineiro I
 (recupero um texto publicado em 2004 como consequência da vinda de Niles Eldredge à Gulbenkian, no próximo dia 13 de Fevereiro, para uma palestra integrada na comemoração dos 150 anos da publicação da Origem das espécies de Charles Darwin)
(recupero um texto publicado em 2004 como consequência da vinda de Niles Eldredge à Gulbenkian, no próximo dia 13 de Fevereiro, para uma palestra integrada na comemoração dos 150 anos da publicação da Origem das espécies de Charles Darwin)
 Desde o primeiro dia em que cheguei ao AMNH, para desenvolver a minha investigação em dinossáurios, tive o título desse livro a martelar-me na cabeça.
Desde o primeiro dia em que cheguei ao AMNH, para desenvolver a minha investigação em dinossáurios, tive o título desse livro a martelar-me na cabeça.  Cretácico/Terciário (65 milhões de anos – 75% de fauna e flora, entre os quais todos os dinossáurios não-avianos e amonites). Esta é uma das extinções melhor estudadas e conhecidas, estando na sua génese o bem conhecido impacto de um meteorito que poderá ter tido como aliados fenómenos de vulcanismo intenso.
Cretácico/Terciário (65 milhões de anos – 75% de fauna e flora, entre os quais todos os dinossáurios não-avianos e amonites). Esta é uma das extinções melhor estudadas e conhecidas, estando na sua génese o bem conhecido impacto de um meteorito que poderá ter tido como aliados fenómenos de vulcanismo intenso.
Patas para que vos quero!
![]()
 A propósito do aparecimento dos primeiros tetrápodes e do Tiktaalik, republico um post e artigo de jornal que publiquei em 2007.
A propósito do aparecimento dos primeiros tetrápodes e do Tiktaalik, republico um post e artigo de jornal que publiquei em 2007.
“Tira daí as patas!”, grita um qualquer mamífero de forma semi-agressiva.
“Dois belos pés!” afirma o comentador desportivo, numa tarde de futebol.
Nós temos. Os peixes não.
A “simples” diferença na forma do esqueleto, como ter “mãos” e “pés” ou autópodes, carrega uma importante história evolutiva desde os peixes até aos animais como nós.
Ao segurar um jornal, o leitor está, em termos evolutivos, a utilizar uma barbatana muito complexa e evoluída, e pertence a um grupo de vertebrados chamados tetrápodes, animais com quatro membros, que incluem animais como os mamíferos, aves, répteis e anfíbios.
O aparecimento dos ossos dos dedos em alguns anfíbios deveria ser resultado de nova “maquinaria” genética, pois todas as estruturas orgânicas são o resultado da informação que está contida nos genes. Será assim?
Um estudo publicado, a 24 de Maio (de 2007), na revista Nature, refere que os genes necessários à formação dos dedos das “mãos” e “pés” dos tetrápodes têm uma história que remonta há 360 milhões de anos ou seja antes de os animais terem feito a “invasão” da terra. O estudo molecular dos genes HoxD (genes reguladores do desenvolvimento em diferentes organismos e áreas do corpo, concretamente no desenvolvimento do esqueleto apendicular, i.e., dos membros) vem mostrar que o património genético necessário já estava presente em peixes primitivos como o actual peixe actinopterígeo Polyodon spathula, considerado um autêntico fóssil vivo.
A análise genética deste animal permitiu afinar as informações paleontológicas com as da biologia do desenvolvimento, possibilitando que estas analisassem dados genéticos de peixes menos “evoluídos” – os actinopterígeos – e os comparassem com os dos tetrápodes. Tradicionalmente, estas análises eram efectuadas em peixes mais “evoluídos”, os teleósteos.
Os estudos paleontológicos em exemplares de transição morfológica entre peixes e animais com verdadeiros membros locomotores deixavam em aberto a possibilidade daquela “revolução” evolutiva se ter dado de uma forma rápida em termos de tempo geológico mas o Polyodon revelou que o património genético que permitiu o aparecimento de verdadeiras patas é mais antigo do que se suponha.
Fundamental para se compreender esta “novela” científica é o conhecimento dos fósseis de transição deste trajecto evolutivo.
Os “fotogramas” que permitem visualizar as alterações morfológicas entre as barbatanas e verdadeiros membros locomotores são vários. Conhecia-se já há algum tempo a parte mais inicial do “filme” – os peixes Eusthenopteron e Panderichthys – e a mais avançada – os anfíbios do Devónico superior como Acanthostega e Ichthyostega. Recentemente foi descoberto mais um “fotograma” – o peixe Tiktaalik; este, apresenta um mosaico de características morfológicas antigas e modernas, no trajecto evolutivo para o aparecimento de verdadeiros autópodes.
Algumas curiosidades morfológicas destes “primos” afastados: Ichtyostega possuía sete dedos em cada pata; o Acanthostega, oito. Desculpem mas não resisto a dizer: “vão-se as barbatanas mas fiquem os dedos!”.
Da próxima vez que um qualquer criacionista falar em falta de fósseis de transição nada como descrever estes belos nomes – Eusthenopteron, Panderichthys, Acanthostega, Tiktaalik e Ichthyostega!
Um outro estudo, de 2006 e desta vez embriológico, levado a cabo em Barcelona, permitiu analisar o processo de formação e disposição de dois ossos do pé em embriões humanos – o calcâneo (osso que constitui o nosso calcanhar) e o astrágalo, ambos ossos do pé.
Foram descritas semelhanças morfológicas entre um embrião humano de 33 dias, nas extremidades inferiores, com barbatanas; aos 54 dias o calcâneo e o astrágalo estão localizados no mesmo preciso local que em Bauria cynops, um réptil mamaliforme que viveu há 260 milhões de anos. As semelhanças anatómicas de posicionamento às 8 semanas e meia dos ossos referidos são enormes entre o embrião humano e a espécie fóssil Diademodon, que viveu há 230 milhões de anos.
Os autores afirmam que, nesta fase, o posicionamento, e respectivas consequências ao nível da locomoção, dos ossos analisados estão a meio “caminho” entre répteis e mamíferos. Este tipo de análises incr
ementa o conhecimento morfológico efectuado por vários autores no séc. XIX, mesmo antes de Darwin publicar a sua obra magna, como Karl Ernst von Baer (1792-1876), que notou semelhanças morfológicas entre embriões de grupos diferentes.
Conta a “tradição”, que von Baer, trabalhava no seu gabinete, e encontrou dois frascos com embriões de aves e lagartos; sem rótulos, não os pôde distinguir à primeira vista…
 Von Baer propôs que estádios embrionário iniciais conservavam padrões morfológicos comuns a vária espécies sendo os estádios mais avançados reveladores de divergência morfológica – as similitudes observadas entre embriões humanos e espécies do passado comprovam que anda bem que os frascos de von Baer deveriam ter os rótulos!
Von Baer propôs que estádios embrionário iniciais conservavam padrões morfológicos comuns a vária espécies sendo os estádios mais avançados reveladores de divergência morfológica – as similitudes observadas entre embriões humanos e espécies do passado comprovam que anda bem que os frascos de von Baer deveriam ter os rótulos!Resumindo: espécies que divergem morfologicamente em estádios mais iniciais irão ser morfologicamente mais distintas em estádios adultos.
Von Baer foi pioneiro nas propostas que fez ao nível do desenvolvimento embrionário sendo o seu trabalho basilar numa das áreas mais importantes das Biologia actual – a evolução e o desenvolvimento, Evo-Devo.
De tudo o que vimos só me resta afirmar que a as teorias evolutivas que explicam o nosso trajecto na história da Terra têm cada vez mais “pés para andar…!”
(Publicado no jornal O Primeiro de Janeiro a 31/5/2007)
BIBLIOGRAFIA
Carroll, R.L., Irwin J. & Green, D.M. 2005. Thermal physiology and the origin of terrestriality in vertebrates. Zool. J. Linn. Soc. 143: 345-358.
Carroll, S. B. 2005. Endless Forms Most Beautiful: The New Science of Evo Devo and the Making of the Animal Kingdom, W. W. Norton & Company.
Davis MC, Dahn RD, & Shubin NH (2007). An autopodial-like pattern of Hox expression in the fins of a basal actinopterygian fish. Nature, 447 (7143), 473-6 PMID: 17522683
Evo-Devo – http://www.pnas.org/cgi/content/full/97/9/4424
Goodwin, B. 1994. How the Leopard Changed its Spots, Phoenix Giants.
Isidro, A. & Vazquez, M.T. 2006. Phylogenetic and ontogenetic parallelisms on talo-calcaneal superposition. The Foot 16, 1-15.
FIGURAS
Carroll, R.L., Irwin J. & Green, D.M. 2005. Thermal physiology and the origin of terrestriality in vertebrates. Zool. J. Linn. Soc. 143: 345-358.
Clack, J. 2002. An early tetrapod from Romer’s Gap, Nature 418, 72 – 76.
Horder, T.J. 2006. Gavin Rylands de Beer: how embryology foreshadowed the dilemmas of the genome. Nat Rev Genet. 7(11):892-8.
Deixar de ser ilha*
 Traços e traços.
Traços e traços.
Linhas que unem tipos e formas.
As ilhas imutáveis e isoladas que eram as espécies, criaram laços.
As mais próximas e as mais afastadas, todas unidas pelas pontes de Darwin.
Imagem* – a primeira árvore evolutiva desenhada por Darwin, em 1837. A única ilustração de “A Origem das Espécies” é uma árvore evolutiva.
Referência – Gregory, T.R. 2008. Understanding Evolutionary Trees. Evolution Education Outreach 1: 121-137.


{kind=link}