


Falsificações naturais
 Os Executivos são todos iguais.
Os Executivos são todos iguais.
Estejamos onde estivermos, conseguimos identificar um quadro importante de uma empresa – o vestuário, o calçado e os adereços são semelhantes. Independentemente das diferentes empresas a que pertencem, percursos de vida, educação e anos de carreira, um executivo transmite uma imagem perfeitamente identificável, estejamos em Nova Iorque, Tóquio ou na Bolsa de Lisboa.
Mas para que utilizo eu uma das imagens de marca do capitalismo?
Tal como os executivos adoptam uma imagem semelhante, a Natureza reproduz formas e funções semelhantes em organismos muito diferentes.
Tubarões e golfinhos, ao nível do seu plano corporal, são muito semelhantes, apesar de um ser um peixe e o outro um mamífero, e estarem separados evolutivamente por 400 milhões de anos.
Tubarões e golfinhos podem ser apontados como exemplos de Evolução Convergente – aquisição independente de características físicas semelhantes por parte de seres vivos muito diferentes.
 Existem outros exemplos de Evolução Convergente: os membros anteriores das aves, dos morcegos e dos pterossáurios (répteis voadores, parentes e contemporâneos dos dinossáurios) apresentam formas seme lhantes. Embora de grupos diferentes, separados por milhões de anos de evolução, possuem estruturas anatómicas que lhes permitem (ou permitiam, no caso dos pterossáurios) uma mesma função: voar. A locomoção bípede (somente nos dois membros posteriores) evoluiu de forma convergente nos humanos e nas aves – nestas surgiu há mais de 200 milhões de anos, nos seus dinossáurios antepassados; nos seres humanos, a transição para a locomoção bípede, há uns meros milhões de anos…
Existem outros exemplos de Evolução Convergente: os membros anteriores das aves, dos morcegos e dos pterossáurios (répteis voadores, parentes e contemporâneos dos dinossáurios) apresentam formas seme lhantes. Embora de grupos diferentes, separados por milhões de anos de evolução, possuem estruturas anatómicas que lhes permitem (ou permitiam, no caso dos pterossáurios) uma mesma função: voar. A locomoção bípede (somente nos dois membros posteriores) evoluiu de forma convergente nos humanos e nas aves – nestas surgiu há mais de 200 milhões de anos, nos seus dinossáurios antepassados; nos seres humanos, a transição para a locomoção bípede, há uns meros milhões de anos…
Os leitores e os pombos partilham, convergentemente, este tipo particular de locomoção com alguns roedores, com os cangurus e com alguns lagartos – facultativamente nestes.
Mas qual o “motivo” da Natureza para organismos tão afastados, em termos evolutivos, apresentem estruturas e funções tão semelhantes?
A primeira justificação passa pelos condicionamentos de design e de eficiência orgânica.
Quem não tem muita paciência para cuidar de plantas em casa conhece os cactos. Estas plantas apresentam formas características, adaptadas aos climas desérticos – forma alongada ou arredondada, sem folhas (para não perderem água) e as folhas que possuem estão transformadas em espinhos e tecidos internos capazes de retenção de água. Mas, onde ia eu? A maioria dos amantes de cactos não sabe é que… está a ser “traída”! Não possuem cactos (família Cactaceae e originários da América do Norte) mas sim plantas da família Euphorbiaceae. Mas a “traição” é justificável… As plantas que consideramos cactos desenvolveram, pelo fenómeno de evolução convergente, formas idênticas às plantas dos westerns. Vivendo em climas igualmente áridos, mas em África, as Euphorbiaceae necessitaram de adaptar a sua estrutura para evitar perdas de água e…”imitaram” os cactos!
A maioria dos amantes de cactos não sabe é que… está a ser “traída”! Não possuem cactos (família Cactaceae e originários da América do Norte) mas sim plantas da família Euphorbiaceae. Mas a “traição” é justificável… As plantas que consideramos cactos desenvolveram, pelo fenómeno de evolução convergente, formas idênticas às plantas dos westerns. Vivendo em climas igualmente áridos, mas em África, as Euphorbiaceae necessitaram de adaptar a sua estrutura para evitar perdas de água e…”imitaram” os cactos!
A segunda razão, por detrás da evolução convergente, envolve o sucesso da imitação.
Podemos utilizar um exemplo da cultura humana – a música. Já todos nós constatámos que quando um grupo musical tem sucesso logo aparecem vários outros a o imitar. As condições do mercado e do gosto musical num determinado momento são as certas, de maneira que os imitadores também vingam, por se aproveitarem de um modelo vencedor.
Na Natureza, a “imitação” estrutural também funciona de maneira equivalente. Por exemplo quer os veados quer os cavalos desenvolveram membros finos e esguios, assentes no desenvolvimento do dedo III (central) e redução dos restantes. Estas adaptações permitiam a optimização da corrida. Quer uns quer outros tinham o mesmo tipo de predadores e, ao longo de milhões, desenvolveram anatomias semelhantes.
Estas adaptações permitiam a optimização da corrida. Quer uns quer outros tinham o mesmo tipo de predadores e, ao longo de milhões, desenvolveram anatomias semelhantes.
A Selecção Natural conduziu espécies competidoras de um mesmo ecossistema por trilhos evolutivos paralelos, pois o sucesso evolutivo estava dependente da economia da forma, da função e do design. Todos nós já constatámos que quando um determinado produto tem sucesso comercial, logo aparecem imitações.
É o ritmo da sociedade de consumo.
Ainda bem que não existem patentes na Natureza!
(Publicado no jornal O Primeiro de Janeiro a 16/02/2006)
A Moby-Dick pode esperar…eis o Indohyus!
 A maioria das pessoas desconhece que os cetáceos, grupo a que pertencem as baleias e golfinhos, já foram animais terrestres.
A maioria das pessoas desconhece que os cetáceos, grupo a que pertencem as baleias e golfinhos, já foram animais terrestres.
Na sua história evolutiva verificaram-se alterações morfológicas que lhes permitiram um “regresso ao mar”.
Uma das características deste grupo é serem, assim, totalmente aquáticos. Para além deste factor são os maiores animais que já existiram – a baleia-azul, com um máximo na 33 m de comprimento e 190 000 kg de peso, mas podendo ter “apenas” 1,4 m e 45 kg, como a Toninha da Califórnia ou vaquita (Phocoena sinus).
Estes dois extremos do grupo Cetacea colocam várias questões evolutivas importantes, e algumas semelhante às colocadas nos dinossáurios saurópodes: que modificações sofreram estes animais para atingirem tamanhos descomunais? E como se deram esses processos?
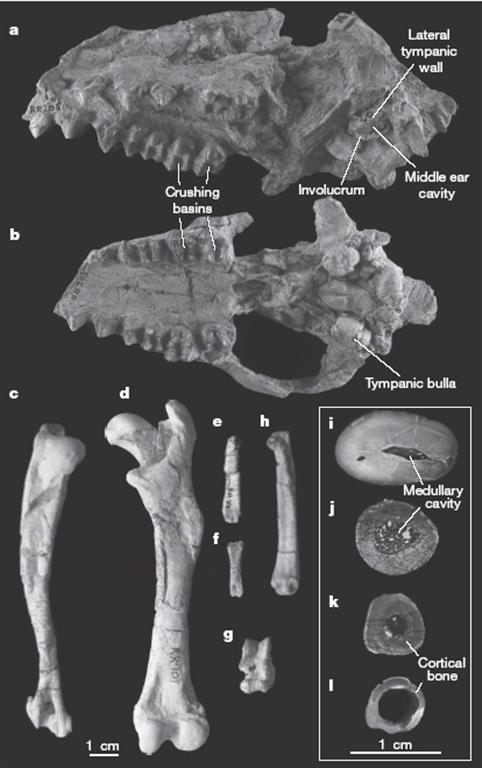
Entre as alterações morfológicas verificadas na evolução dos cetáceos contam-se a redução do esqueleto apendicular, a alteração da forma dos dentes e modificações na estrutura do ouvido interno.
 UM NOVO ELEMENTO NA HISTÓRIA DOS CETACEA
UM NOVO ELEMENTO NA HISTÓRIA DOS CETACEA
Depois de já anteriormente ter levantado a ponta do véu sobre a história evolutiva dos Cetacea, foi publicado hoje, na revista Nature.
Uma das conclusões deste estudo é o da proximidade de parentesco e semelhanças morfológicas entre o Indohyus (família Raoellidae, pertencente á ordem Artiodactyla) e os cetáceos. Esta descoberta permite inferir que o habitat aquático terá entrado na vida destes animais antes mesmo de surgirem os verdadeiros Cetacea. Este estudo aponta também que a mudança de dieta terá surgido na “transição” dos Artiodactyla para os Cetacea.
INDOHYUS
Outra das questões ainda não totalmente esclarecidas diz respeito à “causa” evolutiva que explique a transição do meio terrestre para o meio aquático destes animais. Alguns autores referem a dieta como sendo o fio condutor dessa “viagem”.
As evidências morfológicas surgem através dos dentes fossilizados deste grupo, que apesar de serem perfeitamente diferenciáveis das espécies actuais não permitem inferir com completo rigor a dieta do animal.

ESMALTE E DIETA
Com o objectivo de averigua o carácter aquático do Indohyus, este novo estudo incorpora a análise da proporção entre os isótopos δ18O e δ13C do esmalte dentário. Estes isótopos são bastante estáveis após a morte do animal e posterior conjunto de fenómenos conducentes à sua fossilização e podem ser, e são, utilizados como um indicador do tipo de dieta do animal em estudo. Por exemplo, o isótopo δ18 do oxigénio revela quer a alimentação quer o tipo de água ingeridas, tendo-se verificado que os valores de δ18 presentes no esmalte do Indohyus eram inferiores aos dos mamíferos quer terrestres quer semi-aquáticos, do Eocénico.
Este facto permite inferir que este animal viveria num ambiente aquático, embora não se podendo afirmar se exclusivamente.
Apesar de passar muito tempo dentro de água, alimentar-se-ia também de vegetação em terra, um pouco à semelhança do que acontece com o hipopótamo.
A análise morfológica dos ossos encontrados e da composição química do esmalte dentário permite aos paleontólogos afirmar que o Indohyus não era um nadador exímio, tendo provavelmente vivido em ambiente aquáticos de pequena profundidade, com os membros assentes ou semi-assentes no fundo. Este animal alimentava-se também em terra, embora este estudo aponte a possibilidade de uma dieta aquática.
TIPOS LOCOMOÇÂO AQUÁTICA DOS “VELHOS”
CETACEA
Se os modernos cetáceos apresentam formas muito semelhantes de locomoção aquática, o mesmo não se pode afirmar dos seus directos antepassados directos. No Eocénico (entre os 55 e os 34 milhões de anos atrás) os cetáceos apresentavam diversas morfologias corporais e consequentes modos distintos de natação que iam do balanço da barbatana caudal (nos Dorudontidae, semelhantes a golfinhos) até ao simples “remar” com os quatro membros (nos Pakicetidae).
GOULD
Stephen Jay Gould descreveu grande parte das “peripécias” paleo-cetáceas no seu ensaio mensal na revista do American Museum of Natural History “Natural History”, em 1994. O artigo “Hooking Leviathan by Its Past”, foi compilado no livro “Dinosaur in a Haystack”, editado em Portugal pela Gradiva, mas não me recordo do título…
REFERÊNCIAS
Gingerich PD, Arif M, Bhatti MA, Anwar M, Sanders WJ. 1997. Basilosaurus drazindai and Basiloterus hussaini, new Archaeoceti (Mammalia, Cetacea) from the middle Eocene Drazinda Formation, with revised interpretation of ages of whale-bearing strata in the Kirthar Group of the Sulaiman Range, Punjab (Pakistan). Contrib. Mus. Paleontol. Univ. Mich. 30:55-81
Gingerich PD, Haq M, Zalmout IS, Khan IH, Malkani MS. 2001. Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science 293:2239-42
Gingerich PD, Raza SM, Arif M, Anwar M, Zhou X. 1994. New whale from the Eocene of Pakistan and the origin of cetacean swimming. Nature 368:844-47
Gingerich PD, Smith BH, Simons AL. 1990. Hind limbs of Eocene Basilosaurus: evidence of feet in whales. Science 249:154-57
Gingerich PD, Wells NA, Russell DE, Shah SMI. 1983. Origin of whales in epicontinental remnant seas: newevidence from the early Eocene of Pakistan. Science 220:403-6
Thewissen, J. G. M. & Williams, E. M. 2002. THE EARLY RADIATIONS OF CETACEA (MAMMALIA): Evolutionary Pattern and Developmental Correlations. Annu. Rev. Ecol. Syst. 2002. 33:73-90
Thewissen, J. G., L. N. Cooper, M. T. Clementz, Sunil Bajpai, and B. N. Tiwari. Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nature 450: 1190-1194.
IMAGENS – Carl Buell; Thewissen, J. G. M. et al. 2007; Thewissen, J. G. M. & Williams, E. M. 2002; Thewissen, J. G. M. et al. 2007.
VIDEO
Bioformas – Peek-A-Boo
 De quem é este embrião, na fase “Peek-A-Boo“?
De quem é este embrião, na fase “Peek-A-Boo“?
Curioso o nome dado – “The distal extremities of the forelimbs
overlap, obscuring the face completely and giving this stage its name.”
A recordação que tenho é adolescente e de outro “Peek-A-Boo”…
Chernes e ornitorrincos
 (Publicado no jornal O Primeiro de Janeiro a 18/10/2007)
(Publicado no jornal O Primeiro de Janeiro a 18/10/2007)
“A prova de que Deus tem sentido de humor é o ornitorrinco”, Woody Allen
A atribuição de características humanas a seres vivos ou a elementos naturais – antropomorfismo – é recorrente na literatura, pintura ou na linguagem do dia-a-dia. O inverso – zoomorfismo – que é designar ou conceder singularidades animais a pessoas ou instituições humanas, é frequente na vida política portuguesa. O mais recente caso compara a situação do maior partido da posição ao ornitorrinco – Ornithorhynchus anatinus.
Mas quais os motivos pelos quais o ornitorrinco exerce um fascínio tão grande?
Apesar de mamífero, as fêmeas ornitorrinco colocam ovos sendo as crias posteriormente alimentadas com o leite materno. Este animal apresenta ainda a mandíbula semelhante a um bico de pato, morfologia ímpar entre os mamíferos, morfologia que está na justificação etimológica do seu nome científico Ornithorhynchus – focinho de ave. É esta amálgama de particularidades reptilianas, avianas e mamiferóides que contribuem para que este monotrémato – grupo de mamíferos primitivos a que pertence o ornitorrinco – desempenhe um papel quase mitológico no imaginário colectivo.
É esta amálgama de particularidades reptilianas, avianas e mamiferóides que contribuem para que este monotrémato – grupo de mamíferos primitivos a que pertence o ornitorrinco – desempenhe um papel quase mitológico no imaginário colectivo.
Num dos capítulos de “A Feira dos Dinossáurios” do paleontólogo e historiador da Ciência Stephen Jay Gould, é feita a resenha histórica de como os naturalistas dos sécs. XVIII e XIX observavam o ornitorrinco. Primitivo, ineficiente e imperfeito foram alguns dos adjectivos utilizados então para descrever o mamífero australiano. A Natureza, para aqueles cientistas, deveria apresentar divisões claras e inequívocas na sua diversidade de formas. Estas divisões seriam o resultado da sabedoria divina.
A miscelânea morfológica do ornitorrinco originava, assim, acesos debates não só biológicos mas também teológicos.
 Os monotrématos (como o ornitorrinco) divergiram evolutivamente das linhagens de mamíferos marsupiais (como o canguru) e placentários (como o ser humano) há mais de 100 milhões de anos. Assim, sempre se pensou que estariam desprovidos da fase REM do sono (nos humanos a fase do sono em que se sonha) pois esta seria uma característica moderna. Um estudo de 1998 veio desmentir aquela suposição – o ornitorrinco apresenta sono REM. A quem interessar…
Os monotrématos (como o ornitorrinco) divergiram evolutivamente das linhagens de mamíferos marsupiais (como o canguru) e placentários (como o ser humano) há mais de 100 milhões de anos. Assim, sempre se pensou que estariam desprovidos da fase REM do sono (nos humanos a fase do sono em que se sonha) pois esta seria uma característica moderna. Um estudo de 1998 veio desmentir aquela suposição – o ornitorrinco apresenta sono REM. A quem interessar…O aparente paradoxo, seja político, morfológico ou outro, materializado na forma do ornitorrinco, assenta na errada premissa evolutiva de que os organismos não devem apresentar fusão de características – um animal não deveria colocar ovos e alimentar as suas crias com leite produzido por glândulas mamárias, entre outras.
Gould contrapõe que é precisamente essa amálgama de especificidades que teriam concedido ao ornitorrinco vantagens evolutivas.
Um outro caso de zoomorfismo político envolveu, no passado recente, um conhecido político português e o poema de Alexandre O’Neill:
Sigamos o cherne, minha amiga!
Desçamos ao fundo do desejo
Atrás de muito mais que a fantasia
E aceitemos, até do cherne um beijo,
Senão já com amor, com alegria…
Adivinhem quem é…
Imagens (fontes) – links nas imagens
“Monstros Marinhos” e rigor científico
 “Não perca esta oportunidade de conhecer répteis marinhos ancestrais e as mais recentes teorias sobre as causas que terão provocado a sua extinção!
“Não perca esta oportunidade de conhecer répteis marinhos ancestrais e as mais recentes teorias sobre as causas que terão provocado a sua extinção!
A exposição está dividida em quatro cenários distintos que apresentam a Terra, em termos geológicos, enquanto planeta em constante mudança e os seus mecanismos de evolução.”
Decidi efectuar esta análise a esta exposição após uma visita informal durante a qual constatei variados erros/omissões/incorrecções científicas.
A análise e revisão feitas estão sistematizadas em seguida.
A exposição é constituída por diversos painéis com breve descrição de criaturas marinhas – a maioria extinta – de grande tamanho: o principal critério unificador.
Não existem fósseis ou réplicas dos exemplares apresentados, apesar de estes serem, na sua maioria, unicamente conhecidos pelo registo fóssil.
Uma das ideias mais promissoras e com “valor de mercado” é a intitulada Cultura Pop – Percepções Culturais e a sua relação com conceitos científicos, neste caso num contexto de criaturas marinhas.
Este conceito, apesar de promissor, não foi suficientemente aproveitado, sendo a sua abordagem limitada a um painel em que é referida, introdutoriamente, a influência daquelas criaturas no imaginário colectivo.
De realçar a excelente qualidade gráfica das ilustrações, sempre com escala humana, que muito contribuem para o aspecto geral, agradável e apelativo.
 Revisão/Análise científica
Revisão/Análise científica
Descrevo seguidamente, de forma que não pretende ser exaustiva, algumas das várias incorrecções/omissões/falhas científicas detectadas, documentadas por fotos.
– Critérios pouco uniformes na designação científica dos exemplares, por exemplo: nomes de grupos genéricos (“Notossauro”, “Ictiossauro”) misturados com espécies, uma vezes identificadas (Thalassomedon haningtoni) outras vezes não, apenas pelo género (Dakosaurus, Henodus, Platypterigius).
– Na parte final (em termos do movimento do público) é apresentada num painel representativo, de forma resumida e cronológica, a história, quer geológica, quer biológica, da Terra. Este painel poderia explorar os intervalos temporais relativos de cada uma das fases da História da Terra, ou seja, cada um dos sub-painéis poderia ter um tamanho proporcional à sua amplitude temporal.
– Estando os períodos temporalmente mais próximos do presente antropomorficamente sobreavaliados, poderia ter-se feito um destaque, com painel isolado, revelando pormenores biológicos/geológicos destes períodos.
– “primeiros tetrapódios” – deveria ser primeiros tetrápodes ou Tetrapoda
– “Pangea” e “Pangeia” – utilizados de forma não coerente: deveria ter-se utilizado Pangeia.
– Thalassomedon haningtoni, assim designada em inglês, surge como Thalassomedon hanington em português, o que dá a sensação de que o nome da espécie se altera do português para inglês.
– “Plioceno” em vez de Pliocénico – todas as referências deveriam ser Pliocénico (período geológico entre os 5.3 e 1.8 milhões de anos).
– “Paleoceno” em vez de Paleocénico – todas as referências deveriam ser Paleocénico (período geológico entre os 65 e 55 milhões de anos).
– “Triássico” em vez de Triásico – todas as referências deveriam ser Triásico (período geológico entre os 251 e 200 milhões de anos).
– “Carbonífero” em vez de Carbónico – todas as referências deveriam ser Carbónico (período geológico entre os 359 e 299 milhões de anos). – Dunkleosteus, mencionado como primeiro animal com reprodução sexuada e com comportamento canibal.
– Dunkleosteus, mencionado como primeiro animal com reprodução sexuada e com comportamento canibal.
Em relação ao facto de ser canibal, faltaria acrescentar a informação de que este comportamento foi inferido a partir de marcas de mandíbulas encontradas num crâneo de Dunkleosteus. Uma vez que este animal seria o maior predador da época, os paleontólogos deduziram que só outro elemento daquela espécie poderia ter infligido tal marca – faltaria adicionar, de forma breve, esta inferência paleontológica.
No que diz respeito à afirmação de que seria o primeiro animal com reprodução sexuada, qualquer pessoa com um mínimo de formação biológica sabe que esta afirmação carece de qualquer sentido. Quereriam os autores referir-se a primeiros animais com dimorfismo sexual?
Uma das ilustrações Dunkleosteus tem como texto de suporte “…comia tudo o que via.” Esta afirmação, apesar de talvez apelativa, parece-me exagerada do ponto de vista biológico, podendo cair facilmente na especulação não-científica.
– “Eric leptocleidus” – esta espécie não existe, tendo sido confundido o nome informal “Eric” dado a um exemplar do género Leptocleidus, descoberto na Austrália.
“Período Câmbrico”
– “…maioritariamente organismos marinhos” – toda a vida existente neste período da Terra era exclusivamente marinha, uma vez que a “invasão” terrestre só aconteceu muito mais tarde.
– A frase, geradora de confusão, é contradita pelo cartaz “Ordovícico”, onde se afirma “vida apenas nos mares…” (ver comentários a esta afirmação em “Período Ordovícico”)
– “Trilobites – fósseis indicadores” – é verdade, mas falta referir de que é que são indicadores – de idade geológica, de ambiente ou de que outro tipo de informação.
– “Myllokunmingia” – faltaria acrescentar o nome completo (Myllokunmingia fengjiaoa, a sua idade (530 milhões de anos) e a proveniência (China, província de Kunming)
“Período Ordovícico”
–“…começa com clima não muito intenso e alta humidade” – a aparente falta de sentido desta frase somente pode ser justificada pela má tradução de “milder” para “não muito intenso”.
–“vida apenas nos mares com, inicialmente, níveis muito altos” – níveis muito altos de quê?
– É representada uma trilobite designada por “Trilobite Gigante”, enquanto um escorpião-marinho já é apresentado como Megalograptus – faltaria a designação científica desta espécie.
“Período Triásico” –“…anfíbios labirintóides” – os anfíbios pertencentes ao grupo Labyrinthodontia têm como designação em português Labirintodontes. Para além disto, é referido que este grupo de anfíbios se extingue no final do Triásico: esta afirmação é incorrecta, uma vez que se conhece pelo menos uma espécie deste grupo – o Koolasuchus cleelandi, do Cretácico inferior da Austrália, prolongando-se, assim, o registo paleontológico dos Labirintodontes (cerca de 55 milhões de anos mais tarde do que referido).
–“…anfíbios labirintóides” – os anfíbios pertencentes ao grupo Labyrinthodontia têm como designação em português Labirintodontes. Para além disto, é referido que este grupo de anfíbios se extingue no final do Triásico: esta afirmação é incorrecta, uma vez que se conhece pelo menos uma espécie deste grupo – o Koolasuchus cleelandi, do Cretácico inferior da Austrália, prolongando-se, assim, o registo paleontológico dos Labirintodontes (cerca de 55 milhões de anos mais tarde do que referido).
–“dinossauros dominam depois da extinção” – até ao final do Triásico, as faunas dominantes eram outros grupos, que não os dinossauros – por exemplo, os grupos Rhynchosauria, répteis herbívoros, os Aetosauria, também herbívoros, e os carnívoros Phytosauria, entre outra fauna.
– traduziu-se “ferns” por abetos, quando deveria ser fetos.
– é referido, no painel deste período, que teria surgido o primeiro tubarão, quando o primeiro representante conhecido do grupo dos tubarões, Doliodus problematicus, data do Devónico inferior (cerca de 200 milhões de anos mais cedo do que referido).
 Conclusão
Conclusão
Esta exposição tem como ponto mais positivo apresentar o tema (raramente abordado no contexto de exposições em Portugal) do registo de vertebrados marinhos mesozóicos – leia-se fauna contemporânea dos dinossáurios mas mas não pertencendo a este grupo.
O ponto mais negativo é a deficiente revisão científica e tradução dos texto dos painéis.
Se se tivesse tido o mesmo cuidado na apresentação e descrição das espécies passadas como o que o Oceanário tem nas espécies presentes, esta exposição poderia constituir um marco idêntico aos das exposições que esta instituição tem levado a cabo no passado.
Faltou-lhe mais formação paleontológica…
P.S.- o Oceanário de Lisboa inaugurou, no passado dia 15 de Junho de 2007, a exposição “Monstros Marinhos”; este texto foi enviado ao Oceanário de Lisboa, a 10 de Julho de 2007, não tendo tido eu, até ao momento, qualquer resposta.
Imagens – links nas imagens
O mar e as entranhas – histórias de bactérias
 (Publicado no jornal O Primeiro de Janeiro a 12/07/2007)
(Publicado no jornal O Primeiro de Janeiro a 12/07/2007)
A escuridão é total. Mas há vida.
Não poderíamos viver nestes ambientes. Mas conhecemo-los.
Ou partes dele.
O fundo do mar.
O nosso sistema digestivo.
O grande desconhecido que é o profundo marinho tem equivalências no invisível interno humano.
O PNAS*, de 5 de Julho de 2007, publicou um estudo sobre a vida microscópica, onde se relacionam os dois ambientes, com personagens que estão mais relacionadas do que até aqui se imaginava.
Quer o sistema digestivo quer o fundo do mar são ambientes inóspitos – escuros e com baixas concentrações de oxigénio.
Ainda assim estão repletos de bactérias.
 Sulfurovum litthotrophicum, descrita em 1984, e Nitratiruptor tergarcus são duas espécies de bactérias do grupo ε-Proteobacteria, que habitam o fundo do mar. Sobrevivem a temperaturas comos as que temos no frigorífico lá de casa, 4º C, até aos 70º C. Vivem ambas no substrato marinho de grandes profundidades obtendo energia através da fixação de azoto provenientes de fontes hidrotermais. São consideradas das mais resistentes formas de vida, pois conseguem sobreviver naqueles ambientes adversos, onde as temperaturas podem atingir mais de 100º C, a profundidades, como no caso da fonte hidrotermal “Menez Gwen” dos Açores, de 1700 m.
Sulfurovum litthotrophicum, descrita em 1984, e Nitratiruptor tergarcus são duas espécies de bactérias do grupo ε-Proteobacteria, que habitam o fundo do mar. Sobrevivem a temperaturas comos as que temos no frigorífico lá de casa, 4º C, até aos 70º C. Vivem ambas no substrato marinho de grandes profundidades obtendo energia através da fixação de azoto provenientes de fontes hidrotermais. São consideradas das mais resistentes formas de vida, pois conseguem sobreviver naqueles ambientes adversos, onde as temperaturas podem atingir mais de 100º C, a profundidades, como no caso da fonte hidrotermal “Menez Gwen” dos Açores, de 1700 m.
Apesar da enorme resistência daquelas bactérias em ambiente natural, só recentemente foram cultivadas em laboratório permitindo que fossem estudadas mais detalhadamente.
O outro grupo de bactérias que ninguém gostaria conhecer, pelo menos na prática ocupa, com maior ou menor frequência, o nosso sistema digestivo.
 A Helicobacter pylori, descoberta em 1982, está presente em metade da população mundial, sendo a causadora da inflamação da mucosa do estômago bem como das úlceras gástricas e do duodeno. A descoberta desta relação concedeu, em 2005, o prémio Nobel da Medicina a Barry Marshall e J. Robin Warren.
A Helicobacter pylori, descoberta em 1982, está presente em metade da população mundial, sendo a causadora da inflamação da mucosa do estômago bem como das úlceras gástricas e do duodeno. A descoberta desta relação concedeu, em 2005, o prémio Nobel da Medicina a Barry Marshall e J. Robin Warren.
O minúsculo ser vivo acompanha a espécie humana desde há muito num fenómeno coevolutivo, facilitado pela sua grande variabilidade genética.
Segundo investigadores do Instituto Max Planck em Berlim, a Helibobacter tem sido transmitido de pais para filhos desde a nossa ancestral saída de África.
Reconstruindo a árvore evolutiva desta bactéria, foi possível identificar dois grandes ramos – um que infecta os europeus e norte-americanos e outro que afecta sobretudo os asiáticos. Essas duas linhagens estão associadas às migrações humanas, permitindo reconstituir essas antigas movimentações.
 Outro dos géneros de bactérias patogénicas analisado foi o Campylobacter jejuni, responsável por intoxicações alimentares nomeadamente a gastroenterite. Os ambientes favoritos para a sua disseminação são leite cru ou mal pasteurizado, aves mal cozinhadas e água não tratada (líquida ou em gelo).
Outro dos géneros de bactérias patogénicas analisado foi o Campylobacter jejuni, responsável por intoxicações alimentares nomeadamente a gastroenterite. Os ambientes favoritos para a sua disseminação são leite cru ou mal pasteurizado, aves mal cozinhadas e água não tratada (líquida ou em gelo).
A equipa de investigadores procedeu à análise do ADN presente nas bactérias que partilham o nosso ambiente digestivo – Helicobacter e Campylobacter – e o das bactérias das profundezas marinhas – Sulfurovum e Nitratiruptor.
Os dois grupos de bactérias apresentaram afinidades genéticas, que lhes possibilitam viverem em ambientes hostis. Entre as semelhanças estão a quase ausência de genes de reparação do ADN. Este facto permite não só a grande adaptação destes seres vivos a novas condições extremas, mas também ao próprio sistema de defesa de um organismo hospedeiro.
 Segundo os investigadores, as bactérias humanas evoluíram a partir de ancestrais de grande profundidade, adquirindo o seu “mau-feitio” quando estabeleceram relações simbióticas com invertebrados marinhos.
Segundo os investigadores, as bactérias humanas evoluíram a partir de ancestrais de grande profundidade, adquirindo o seu “mau-feitio” quando estabeleceram relações simbióticas com invertebrados marinhos.
Não era novidade que tínhamos todos uma origem marinha.
Foi de lá que viemos.
O profundo marinho e o sistema digestivo humano partilham coincidências evolutivas.
O sistema digestivo humano serve de mar a uma variedade de fauna microscópica; sabemos agora que parte dessa fauna tem parentes próximos nos fundos marinhos.
* Proceedings of the National Academy of Sciences
Referências consultadas
-Inagaki, F. et al. 2004. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε -Proteobacteria isolated from Okinawa Trough hydrothermal sediments. International Journal of Systematic and Evolutionary Microbiology, 54, 1477-1482.
-Nakagawa, S. et al. 2007. Deep-sea vent ε-proteobacterial genomes provide insights into emergence of pathogens PNAS published July 5, 2007, 10.1073/pnas.0700687104.
-Nakagawa, S. et al. 2005. Nitratiruptor tergarcus gen. nov., sp. nov. and Nitratifractor salsuginis gen. nov., sp. nov., nitrate-reducing chemolithoautotrophs of the -Proteobacteria isolated from a deep-sea hydrothermal system in the Mid-Okinawa Trough. International Journal of Systematic and Evolutionary Microbiology, 55, 925
Equilíbrio Pontuado

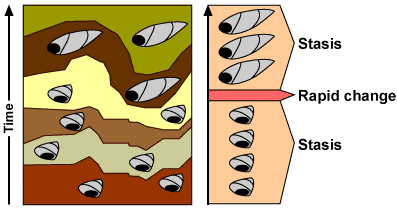
As relações amorosas sofrem do mesmo mecanismo que a Evolução das Espécies – Equilíbrio Pontuado.
Permanecem estáveis e imutáveis durante muito tempo – Estase.
Depois, e em períodos muito curtos, alteram-se e surgem novas relações.
E espécies.
Placentários – e daí?
 O Maelestes gobiensis é um mamífero, descoberto em 1997, escavado no deserto de Gobi em rochas com 75 milhões de anos.
O Maelestes gobiensis é um mamífero, descoberto em 1997, escavado no deserto de Gobi em rochas com 75 milhões de anos.
“E daí?”
O Maelestes gobiensis não é directo ancestral de qualquer grupo de mamíferos modernos apesar de relacionado com os placentários, grupo ao qual pertencemos.
“E daí?”
Das 5416 espécies actuais de mamíferos, 5080 são mamíferos placentários.
“E daí?”
Paleontólogos utilizaram esta espécie como referencial na comparação morfológica de mais de 60 espécies de mamíferos actuais e fósseis.1
“E daí?”
Os mamíferos placentários, das baleias aos ratos, surgiram1, no hemisfério norte, há pouco mais de 65 milhões de anos – quando os dinossáurios se extinguiram.
“E daí?” Este estudo contradiz um outro2, publicado em Março em baseado no ADN de espécies actuais, que concluía que os modernos placentários tinham surgido antes há 100 milhões de anos – antes do desaparecimento dos dinossáurios.
Este estudo contradiz um outro2, publicado em Março em baseado no ADN de espécies actuais, que concluía que os modernos placentários tinham surgido antes há 100 milhões de anos – antes do desaparecimento dos dinossáurios.
“E daí?”
Daí, nada…”apenas” mais uma peça no puzzle na evolução dos mamíferos.
Apenas.
Referências
1 J. R. Wible, G. W. Rougier, M. J. Novacek & R. J. Asher Nature 447, 1003-1006 (21 June 2007)
2 Bininda-Emonds, O.R.P., M. Cardillo, K.E. Jones, R.D.E. MacPhee, R.M.D. Beck, R. Grenyer, S.A. Price, R.A. Vos, J.L. Gittleman, and A. Purvis. 2007. The delayed rise of present-day mammals. Nature 446(7135): 507-512.
Ilustrações: Paul Bowden/CMNH
Foto: John Wible/CMNH
Baleias com patas, a Origem das Espécies e aves fósseis
 (Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
Hoje comemoram-se 146 anos de existência de um livro que teve fortes repercussões pelas suas propostas de Evolução pelo mecanismo de Selecção Natural. A 24 de Novembro de 1859, Charles Darwin publicou “A Origem das Espécies” com uma primeira edição de 1250 exemplares que esgotou no seu dia de lançamento.
Darwin defendia que o meio ambiente e as relações entre os próprios seres vivos exercem uma selecção que favorece os mais aptos enquanto os menos dotados são eliminados, transmitindo-se à geração seguinte as diferenças que facilitam aquela sobrevivência. Ao longo das gerações, essas características acentuam-se e geram uma nova espécie.
Darwin foi convidado em 1831 a participar numa volta ao mundo no navio Beagle promovida pela marinha inglesa. A expedição – que durou cinco anos – tinha o objectivo de melhorar e completar dados cartográficos. Esta viagem foi decisiva para fundamentar as suas teorias evolutivas.
América do Sul, Austrália e Nova Zelândia foram alguns dos locais em que Beagle efectuou paragens. Surpreendeu-se com o grande número de espécies de plantas e de animais que, até então, eram desconhecidos. O que lhe chamou mais atenção foram a enorme diversidade de tentilhões, que só conheceu no arquipélago das Galápagos.
 O registo fóssil
O registo fóssil
Quando Darwin propôs que a Evolução se dava pela Selecção Natural, o registo fóssil oferecia ainda poucas evidências que apoiassem as suas ideias.
Actualmente os paleontólogos contam com mais informação fossilífera do que aquela que dispunha Darwin. Este dedicou dois capítulos da sua “Origem das Espécies” à imperfeição do registo fóssil, provavelmente porque constatava que estaria aí um dos pontos mais fracos da sua argumentação. Dois anos depois da publicação do seu livro, o primeiro exemplar de Archaeopteryx foi descoberto na Baviera, constituindo um dos “elos perdidos” entre as aves e os répteis.
O registo fóssil está longe de ser perfeito – faltam assim muitos elementos de transição na história evolutiva dos seres vivos. Este é um dos argumentos com que os Criacionistas (opositores à Teoria da Evolução, populares sobretudo nos EUA) se valem no seu ataque a Darwin.
As últimas estimativas apontam para que apenas 1% de todas as espécies animais e vegetais que habitaram o nosso planeta ficaram preservadas como fósseis. Razões para esta escassez de informação fóssil são várias, mas podemos apontar que as condições físicas necessárias à preservação de um ser vivo sob a forma de fóssil são muito raras. Grande parte dos seres vivos é constituída por partes moles; este facto contribui igualmente para que o registo fóssil seja desprovido da maioria dos “actores” da vida sobre a Terra.
Mas existem numerosos exemplos que reflectem as etapas de transição entre diversos grupos de organismos, corroborando Darwin.
Para além do já referido Archaeopteryx (descoberto em sedimentos da Baviera com 150 milhões de anos) foi descoberta nos últimos 20 anos toda uma panóplia de formas de transição entre os dinossáurios carnívoros (semelhantes ao Velociraptor) e as actuais aves: Confuciusornis (China, primeira evidência de um bico sem dentes); Iberomesornis (Espanha, apresentava uma estrutura óssea semelhante às aves actuais que permitia inserir a musculatura específica para o voo), entre inúmeros outros exemplos.
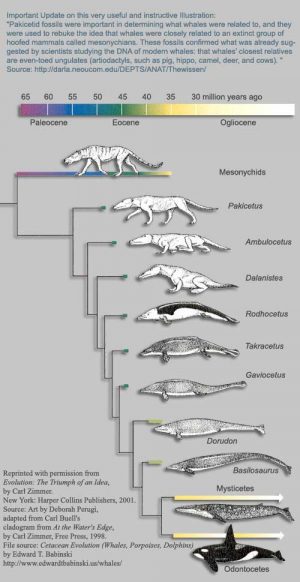 As patas das Baleias
As patas das Baleias
Outro dos exemplos que se podem apresentar para ilustrar as transições evolutivas sofridas pelos organismos é a evolução das baleias.
A maioria das pessoas tem, pelo menos a noção, de que a vida terrestre surgiu a partir de vertebrados que deixaram o ambiente aquático. Todas as formas de animais que ocuparam e ocupam um ambiente terrestre, descendem desses primeiros colonizadores. O que a maioria das pessoas não sabe é que o grupo de animais a que pertencem as actuais baleias descendem de um grupo que “decidiu” voltar a ambientes aquáticos, donde tinham saído há mais 250 milhões de anos.
Em 1983 foram descobertos no Paquistão fósseis de um animal que tinha vivido há cerca de 52 milhões de anos. Este animal, Pakicetus, apresentava ainda corpo com forma para a vida terrestre (membros com capacidade de locomoção em terra) mas tinha um crâneo e dentes com características típicas dos ancestrais dos actuais baleias. Onze anos mais tarde e igualmente no Paquistão foi descoberto o Ambulocetus natans (literalmente baleia caminhante que nada). O Ambulocetus tinha o tamanho de um leão-marinho e apresentava as patas (sim esta baleia ancestral tinha patas!) com capacidades para deslocação em ambiente terrestre. Igualmente exibia os seus pés e mãos com capacidade natatória – ou seja este animal possuía capacidades para se deslocar em ambiente terrestre e aquático.
Aparentemente o Ambulocetus nadava como uma lontra, com movimentos para cima e para baixo (dados da morfologia da sua coluna vertebral atestam-no).
Em 1995 um terceiro elemento de transição foi descoberto – o Dalanistes. Apresentava os membros mais curtos que Ambulocetus, cauda e crâneo mais alongados ou seja mais semelhantes às actuais baleias.
Actualmente mais de uma dúzia de fósseis ilustrativos das transições evolutivas dos cetáceos (grupo a que pertencem as baleias) já foram descobertos.
Complementarmente análises de ADN mitocondrial aos actuais representantes dos cetáceos permitem apontar que estes pertencem ao grupo dos artiodáctilos, mais concretamente são parentes próximos dos hipopótamos.
No dia de aniversário da A Origem das Espécies, que fez com que o Homem descesse de mais um dos seus inúmeros pedestais, podemos afirmar que o registo fóssil é mais um dos motivos de orgulho para Darwin.
Ao contrário do que receava, o trabalho de investigação paleontológico nos últimos 100 anos, permitiu que o registo fóssil seja mais uma prova de que Darwin não estava e não está errado.
Imagem – Werner Horvath: “Charles Robert Darwin – Evolution”. Oil on canvas
A Evolução escondida nos Cartoons
 (Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
A História da vida animal está cheia de alterações na forma dos seus corpos. A análise da forma (análise morfológica) é um das ferramentas no estudo da história evolutiva dos seres vivos.
Um dos essenciais evolucionistas do nosso século – Stephen Jay Gould da Universidade de Harvard – utilizou o melhor da dedução científica para analisar um dos ícones da Disney.
Gould recolheu uma sequência cronológica de Ratos Mickey, desde os anos 20 até à actualidade, analisando alguns traços morfológicos desta figura da banda-desenhada – altura e tamanho geral do crâneo, bem como o tamanho dos olhos. Constatou que ao logo das mais de cinco décadas da história do pequeno ratinho, os parâmetros analisados tinham aumentado. Por outras palavras, Mickey tinha-se transformado: de uma figura de crâneo pequeno e alongado e de olhos pequenos num ratinho de crâneo arredondado e alto com olhos proporcionalmente grandes. Se olharmos para a figura qual dos dois extremos “evolutivos” de Mickey nos parece mais atractivo?
A maioria das pessoas referirá que o da direita (olhos maiores, crâneo mais arredondado). Estas características morfológicas podem ser reconhecidas, na grande maioria dos casos, como sendo características de crias de mamíferos. Todos nós facilmente identificamos que os bebés apresentam um crâneo e olhos proporcionalmente maiores do que os adultos. Estas particularidades têm um efeito sobre os membros de uma determinada espécie e igualmente nos humanos. Características morfológicas de juvenis parecem desencadear reacções de protecção e carinho – um crâneo grande e arredondado, olhos enormes, mandíbula pequena, etc.
Estes são alguns dos traços morfológicos apelativos nos mamíferos. Desta maneira Gould constatou que os desenhadores da Disney alteraram a fisionomia de Mickey atribuindo-lhe características morfológicas juvenis com o objectivo o tornarem emocionalmente mais apelativo.
Pelo contrário, podem ser encontradas algumas morfologias opostas em alguns dos vilões da Disney. Estes apresentam características anatómicas geradoras de desconfiança – crâneo afilado, olhos pequenos, mandíbula proeminente. Enquadram-se neste grupo a Rainha da Branca de Neve, a bruxa da Bela Adormecida e o feiticeiro Jafar em Aladino.
Mas o que tem isto a ver com a evolução?
Gould não perdeu o seu tempo com esta actividade aparentemente estéril. Pretendia ilustrar, com exemplos facilmente reconhecíveis, alguns conceitos da morfologia evolutiva. Uma das noções subjacentes ao estudo dos Mickeys é o da Pedomorfose – aquisição no adulto de um grupo descendente de características morfológicas juvenis do grupo antepassado.
 Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Este conceito está englobado num conjunto maior que é a Heterocronia – em sentido geral, a variação no tempo de aparecimento de uma característica morfológica de um grupo descendente comparativamente ao aparecimento dessa característica no grupo antepassado.
De uma maneira mais simples: podemos ver a “quantidade de transformação” que um indivíduo sofre ao longo do seu desenvolvimento é a mesma, maior ou menor do que do seu antepassado (aqui em sentido de evolutivo e não de progenitor).
O estudo da Heterocronia tem chamado a atenção nos últimos anos dos investigadores da Evolução. É um campo complexo mas muito apelativo e em que têm sido feitas inúmeras descobertas no estudo da morfologia de seres vivos actuais ou dos seus antepassados fósseis.
Outro dos exemplos de Pedomorfose no mundo não-natural é a evolução do símbolo da Shell. O seu logotipo é o bivalve Pecten tendo este “evoluído” desde o início do século por redução do número de “linhas” na concha, aparecendo assim menos complexo e mais juvenil.
Mas existem muito exemplos que podem ilustrar os conceitos evolutivos já referidos: é fazer uma busca na Internet pela “evolução” do Snoopy…



{kind=link}