


Sornas e viagens espaciais
 (Publicado no jornal O Primeiro de Janeiro a 01/11/2007)
(Publicado no jornal O Primeiro de Janeiro a 01/11/2007)
Apesar de muito desejada, a hibernação não passa de um desejo inatingível pelos humanos. Já demos por nós, várias vezes e em alturas de maior cansaço, a cobiçar dormir por vários dias. Desligar e apenas descansar. Passar pelas brasas de forma longa e continuada.
Mas a hibernação é muito mais do que um simples dormir.
Pode definir-se como um estado em que o animal tem uma substancial redução quer na temperatura corporal quer nos gastos energéticos bem como na taxa cardíaca. A redução térmica verificada em animais que hibernam pode atingir mínimos de 5ºC – temperatura semelhante ao interior de um frigorífico, na zona dos vegetais.
A redução de gastos energéticos nos animais que hibernam pode atingir o 1% do habitual embora o mais habitual sejam reduções energéticas para níveis dos 10-20% do normal.
Era tão bom poder, da mesma forma, entrar em “hibernação” de despesas …
Os animais que utilizam esta estratégia fisiológica fazem-no com diversos objectivos: sobreviver em locais que apresentem Invernos com temperaturas muito baixas e/ou em alturas do ano em que a disponibilidade de alimentos seja baixa.
Previamente à hibernação verifica-se grande consumo alimentar pois os animais pretendem acumular gordura com vista à época de carestia que se avizinha.
Mais uma vez o paralelismo para a sociedade humana poderia ser estabelecido…
Um estudo, a ser publicado em Novembro, refere que um marsupial é capaz de hibernar mais de um ano; concretamente foram 367 dias em letargia fisiológica.
 O Cercartetus nanus alimenta-se de néctar e insectos sendo arborícola, i.e., habita nas árvores, de zonas quentes e húmidas do sudoeste australiano. Ao contrário da maioria de outros mamíferos que hibernam para fazer face a baixas temperaturas, o Cercartetus fá-lo quando a carência alimentar surge pelas imprevisíveis condições climatéricas australianas. Este animal apresenta uma enorme capacidade de armazenamento alimentar, sob a forma de gordura, que lhe proporciona, em qualquer altura do ano, entrar em torpor fisiológico.
O Cercartetus nanus alimenta-se de néctar e insectos sendo arborícola, i.e., habita nas árvores, de zonas quentes e húmidas do sudoeste australiano. Ao contrário da maioria de outros mamíferos que hibernam para fazer face a baixas temperaturas, o Cercartetus fá-lo quando a carência alimentar surge pelas imprevisíveis condições climatéricas australianas. Este animal apresenta uma enorme capacidade de armazenamento alimentar, sob a forma de gordura, que lhe proporciona, em qualquer altura do ano, entrar em torpor fisiológico.Contrapondo-se aos registados 367 dias do Cercartetus já tinha sido observado um período de 342 dias em Zapus princeps, um roedor norte-americano.
Poderá questionar-se o leitor sobre o interesse prático destes recordes de inércia…
Para além do conhecimento da diversidade de estratégias no mundo animal, o estudo dos mecanismos fisiológicos utilizados por estes animais poderá contribuir para que viagens espaciais muito longas (até Marte, por exemplo) sejam feitas pelos astronautas em condições parecidas. A redução da actividade fisiológica possibilitará que a viagem seja feita de maneira mais confortável bem como, factor fundamental em viagens espaciais, reduzir as reservas de alimentos, água e oxigénio necessárias a tão longa viagem.
Para ajudar neste processo investigadores da Universidade da Carolina do Norte descobriram dois genes responsáveis pela regulação, em esquilos, de como os organismos usam as reservas energéticas. Estes genes, apesar de envolvidos noutros processos, estão também presentes nos seres humanos.
 Referências:
Referências:
Andrews, M. T., Squire, T. L., Bowen, C. M. & Rollins, M. B. 1998. Low-temperature carbon utilization is regulated by novel gene activity in the heart of a hibernating mammal. Proc. Natl. Acad. Sci. USA Vol. 95, pp. 8392–8397.
Geiser, F. Yearlong hibernation in a marsupial mammal. 2007. Naturwissenschaften 94:941–944
Geiser, F. 2004. Metabolic rate and body temperature Reduction during hibernation and daily torpor. Annu. Rev. Physiol..66:239-274.
Imagens: primeira imagem Cercartetus nanus – essa e as outras – links nas fotos
Cada vez mais altriciais…*
 Antes, ia aos correios, com 6 ou 7 anos, pagar as contas ou levantar uma encomenda à minha mãe.
Antes, ia aos correios, com 6 ou 7 anos, pagar as contas ou levantar uma encomenda à minha mãe.
Andava de uma lado para o outro, ia a casa dos meus amigos desafiá-los para jogar à bola ou simplesmente para passear.
Agora, sobrinhos com o dobro da idade que eu tinha quando circulava sozinho, têm cada vez menos autonomia.
Vão às discotecas, mas são “deixados e levantados” à porta; passam horas ao telemóvel com os amigos, mas raramente os vêm, fora das câmaras da net; não aparecem lá por casa, marcam-se horas.
Agora, na idade adulta, saímos cada vez mais tarde do tecto dos pais; casamos mais tarde; temos filhos mais tarde;…
Estaremos a ficar cada vez mais altriciais?
Ou as ameaças dos “predadores” cada vez mais intensas?
Ou apenas paranóicos?
 * o grau de desenvolvimento e de dependência das crias em relação aos progenitores pode ser diverso.
* o grau de desenvolvimento e de dependência das crias em relação aos progenitores pode ser diverso.
Basta comparar um vitelo (precocial- bem desenvolvido e ao fim de pouco tempo já se desloca autonomamente) com as crias de coelho (altricial – com um grau de desenvolvimento baixo e completamente dependentes dos progenitores).
Imagens – “Views of a Fetus in the Womb”, Leonardo da Vinci ; e daqui com diversos vídeos e textos.
Pinguim…gigante!
 Julia Clarke ainda deve estar a pensar que tem que adiar o seu livro da história de culinária – uma das muitas ideias, aparentemente absurdas, que trocávamos enquanto fumávamos no telhado do American Museum of Natural History, em Nova Iorque.
Julia Clarke ainda deve estar a pensar que tem que adiar o seu livro da história de culinária – uma das muitas ideias, aparentemente absurdas, que trocávamos enquanto fumávamos no telhado do American Museum of Natural History, em Nova Iorque.
É que a nova espécie de pinguim – Icadyptes salasi – que agora descreveu não lhe deve ter deixado muito tempo livre.
Este novo “cromo” tinha 1,5 m de altura e viveu (há 36 milhões de anos) muito mais a norte do que os seus “primos” contemporâneos – na costa do Peru.
Para além de ser uma nova espécie com um tamanho muito apreciável (apesar de mais pequeno que o Anthropornis nordenskjoeldi , com1,7 m de altura e 90 kg de peso) viveu numa zona desértica, num período da história em que a Terra era bastante mais quente do que é actualmente.
Estas novas aquisições da biologia terrestre provocam que os paleontólogos tenham que rever a história evolutiva dos pinguins – a migração para norte, que se pensava ter ocorrido entre 6 e 8 milhões de anos atrás, iniciou-se há mais tempo.
 O membro mais antigo do grupo dos pinguins viveu, no que é hoje a Nova Zelândia, há 61 milhões de anos.
O membro mais antigo do grupo dos pinguins viveu, no que é hoje a Nova Zelândia, há 61 milhões de anos.
Para além do “gigante” Icadyptes salasi foi descoberta outra nova espécie – Perudyptes devriesi – que era um pouco mais antigo – data de há 42 milhões de anos e de tamanho semelhante ao pinguim-rei.
Apesar de ambas as espécies estarem adaptadas a climas quentes, este facto não significa que alterações climáticas não tenham um impacto negativo nas espécies actuais, refere Julia Clarke ao jornal The Guardian.
Imagem – Icadyptes salasi e Perudyptes devriesi
Fonte – AP
Referências
Edição on-line da revista Proceedings of the National Academy of Sciences, de 25 de Junho de 2007.
Baleias com patas, a Origem das Espécies e aves fósseis
 (Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
Hoje comemoram-se 146 anos de existência de um livro que teve fortes repercussões pelas suas propostas de Evolução pelo mecanismo de Selecção Natural. A 24 de Novembro de 1859, Charles Darwin publicou “A Origem das Espécies” com uma primeira edição de 1250 exemplares que esgotou no seu dia de lançamento.
Darwin defendia que o meio ambiente e as relações entre os próprios seres vivos exercem uma selecção que favorece os mais aptos enquanto os menos dotados são eliminados, transmitindo-se à geração seguinte as diferenças que facilitam aquela sobrevivência. Ao longo das gerações, essas características acentuam-se e geram uma nova espécie.
Darwin foi convidado em 1831 a participar numa volta ao mundo no navio Beagle promovida pela marinha inglesa. A expedição – que durou cinco anos – tinha o objectivo de melhorar e completar dados cartográficos. Esta viagem foi decisiva para fundamentar as suas teorias evolutivas.
América do Sul, Austrália e Nova Zelândia foram alguns dos locais em que Beagle efectuou paragens. Surpreendeu-se com o grande número de espécies de plantas e de animais que, até então, eram desconhecidos. O que lhe chamou mais atenção foram a enorme diversidade de tentilhões, que só conheceu no arquipélago das Galápagos.
 O registo fóssil
O registo fóssil
Quando Darwin propôs que a Evolução se dava pela Selecção Natural, o registo fóssil oferecia ainda poucas evidências que apoiassem as suas ideias.
Actualmente os paleontólogos contam com mais informação fossilífera do que aquela que dispunha Darwin. Este dedicou dois capítulos da sua “Origem das Espécies” à imperfeição do registo fóssil, provavelmente porque constatava que estaria aí um dos pontos mais fracos da sua argumentação. Dois anos depois da publicação do seu livro, o primeiro exemplar de Archaeopteryx foi descoberto na Baviera, constituindo um dos “elos perdidos” entre as aves e os répteis.
O registo fóssil está longe de ser perfeito – faltam assim muitos elementos de transição na história evolutiva dos seres vivos. Este é um dos argumentos com que os Criacionistas (opositores à Teoria da Evolução, populares sobretudo nos EUA) se valem no seu ataque a Darwin.
As últimas estimativas apontam para que apenas 1% de todas as espécies animais e vegetais que habitaram o nosso planeta ficaram preservadas como fósseis. Razões para esta escassez de informação fóssil são várias, mas podemos apontar que as condições físicas necessárias à preservação de um ser vivo sob a forma de fóssil são muito raras. Grande parte dos seres vivos é constituída por partes moles; este facto contribui igualmente para que o registo fóssil seja desprovido da maioria dos “actores” da vida sobre a Terra.
Mas existem numerosos exemplos que reflectem as etapas de transição entre diversos grupos de organismos, corroborando Darwin.
Para além do já referido Archaeopteryx (descoberto em sedimentos da Baviera com 150 milhões de anos) foi descoberta nos últimos 20 anos toda uma panóplia de formas de transição entre os dinossáurios carnívoros (semelhantes ao Velociraptor) e as actuais aves: Confuciusornis (China, primeira evidência de um bico sem dentes); Iberomesornis (Espanha, apresentava uma estrutura óssea semelhante às aves actuais que permitia inserir a musculatura específica para o voo), entre inúmeros outros exemplos.
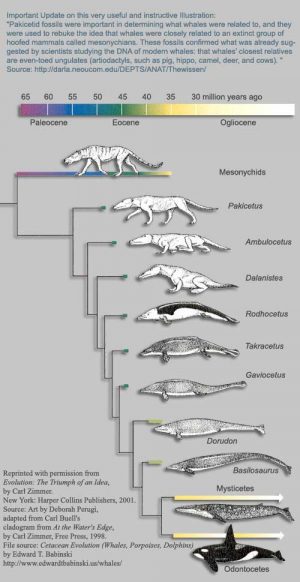 As patas das Baleias
As patas das Baleias
Outro dos exemplos que se podem apresentar para ilustrar as transições evolutivas sofridas pelos organismos é a evolução das baleias.
A maioria das pessoas tem, pelo menos a noção, de que a vida terrestre surgiu a partir de vertebrados que deixaram o ambiente aquático. Todas as formas de animais que ocuparam e ocupam um ambiente terrestre, descendem desses primeiros colonizadores. O que a maioria das pessoas não sabe é que o grupo de animais a que pertencem as actuais baleias descendem de um grupo que “decidiu” voltar a ambientes aquáticos, donde tinham saído há mais 250 milhões de anos.
Em 1983 foram descobertos no Paquistão fósseis de um animal que tinha vivido há cerca de 52 milhões de anos. Este animal, Pakicetus, apresentava ainda corpo com forma para a vida terrestre (membros com capacidade de locomoção em terra) mas tinha um crâneo e dentes com características típicas dos ancestrais dos actuais baleias. Onze anos mais tarde e igualmente no Paquistão foi descoberto o Ambulocetus natans (literalmente baleia caminhante que nada). O Ambulocetus tinha o tamanho de um leão-marinho e apresentava as patas (sim esta baleia ancestral tinha patas!) com capacidades para deslocação em ambiente terrestre. Igualmente exibia os seus pés e mãos com capacidade natatória – ou seja este animal possuía capacidades para se deslocar em ambiente terrestre e aquático.
Aparentemente o Ambulocetus nadava como uma lontra, com movimentos para cima e para baixo (dados da morfologia da sua coluna vertebral atestam-no).
Em 1995 um terceiro elemento de transição foi descoberto – o Dalanistes. Apresentava os membros mais curtos que Ambulocetus, cauda e crâneo mais alongados ou seja mais semelhantes às actuais baleias.
Actualmente mais de uma dúzia de fósseis ilustrativos das transições evolutivas dos cetáceos (grupo a que pertencem as baleias) já foram descobertos.
Complementarmente análises de ADN mitocondrial aos actuais representantes dos cetáceos permitem apontar que estes pertencem ao grupo dos artiodáctilos, mais concretamente são parentes próximos dos hipopótamos.
No dia de aniversário da A Origem das Espécies, que fez com que o Homem descesse de mais um dos seus inúmeros pedestais, podemos afirmar que o registo fóssil é mais um dos motivos de orgulho para Darwin.
Ao contrário do que receava, o trabalho de investigação paleontológico nos últimos 100 anos, permitiu que o registo fóssil seja mais uma prova de que Darwin não estava e não está errado.
Imagem – Werner Horvath: “Charles Robert Darwin – Evolution”. Oil on canvas
A Evolução escondida nos Cartoons
 (Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
A História da vida animal está cheia de alterações na forma dos seus corpos. A análise da forma (análise morfológica) é um das ferramentas no estudo da história evolutiva dos seres vivos.
Um dos essenciais evolucionistas do nosso século – Stephen Jay Gould da Universidade de Harvard – utilizou o melhor da dedução científica para analisar um dos ícones da Disney.
Gould recolheu uma sequência cronológica de Ratos Mickey, desde os anos 20 até à actualidade, analisando alguns traços morfológicos desta figura da banda-desenhada – altura e tamanho geral do crâneo, bem como o tamanho dos olhos. Constatou que ao logo das mais de cinco décadas da história do pequeno ratinho, os parâmetros analisados tinham aumentado. Por outras palavras, Mickey tinha-se transformado: de uma figura de crâneo pequeno e alongado e de olhos pequenos num ratinho de crâneo arredondado e alto com olhos proporcionalmente grandes. Se olharmos para a figura qual dos dois extremos “evolutivos” de Mickey nos parece mais atractivo?
A maioria das pessoas referirá que o da direita (olhos maiores, crâneo mais arredondado). Estas características morfológicas podem ser reconhecidas, na grande maioria dos casos, como sendo características de crias de mamíferos. Todos nós facilmente identificamos que os bebés apresentam um crâneo e olhos proporcionalmente maiores do que os adultos. Estas particularidades têm um efeito sobre os membros de uma determinada espécie e igualmente nos humanos. Características morfológicas de juvenis parecem desencadear reacções de protecção e carinho – um crâneo grande e arredondado, olhos enormes, mandíbula pequena, etc.
Estes são alguns dos traços morfológicos apelativos nos mamíferos. Desta maneira Gould constatou que os desenhadores da Disney alteraram a fisionomia de Mickey atribuindo-lhe características morfológicas juvenis com o objectivo o tornarem emocionalmente mais apelativo.
Pelo contrário, podem ser encontradas algumas morfologias opostas em alguns dos vilões da Disney. Estes apresentam características anatómicas geradoras de desconfiança – crâneo afilado, olhos pequenos, mandíbula proeminente. Enquadram-se neste grupo a Rainha da Branca de Neve, a bruxa da Bela Adormecida e o feiticeiro Jafar em Aladino.
Mas o que tem isto a ver com a evolução?
Gould não perdeu o seu tempo com esta actividade aparentemente estéril. Pretendia ilustrar, com exemplos facilmente reconhecíveis, alguns conceitos da morfologia evolutiva. Uma das noções subjacentes ao estudo dos Mickeys é o da Pedomorfose – aquisição no adulto de um grupo descendente de características morfológicas juvenis do grupo antepassado.
 Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Este conceito está englobado num conjunto maior que é a Heterocronia – em sentido geral, a variação no tempo de aparecimento de uma característica morfológica de um grupo descendente comparativamente ao aparecimento dessa característica no grupo antepassado.
De uma maneira mais simples: podemos ver a “quantidade de transformação” que um indivíduo sofre ao longo do seu desenvolvimento é a mesma, maior ou menor do que do seu antepassado (aqui em sentido de evolutivo e não de progenitor).
O estudo da Heterocronia tem chamado a atenção nos últimos anos dos investigadores da Evolução. É um campo complexo mas muito apelativo e em que têm sido feitas inúmeras descobertas no estudo da morfologia de seres vivos actuais ou dos seus antepassados fósseis.
Outro dos exemplos de Pedomorfose no mundo não-natural é a evolução do símbolo da Shell. O seu logotipo é o bivalve Pecten tendo este “evoluído” desde o início do século por redução do número de “linhas” na concha, aparecendo assim menos complexo e mais juvenil.
Mas existem muito exemplos que podem ilustrar os conceitos evolutivos já referidos: é fazer uma busca na Internet pela “evolução” do Snoopy…

Rápidas e promíscuas
 As chitas (Acinonyx jubatus) constituem um dos animais favoritos dos documentários da vida selvagem.
As chitas (Acinonyx jubatus) constituem um dos animais favoritos dos documentários da vida selvagem.
Graciosas, esguias, tornam-se facilmente em heroínas das novelas da vida selvagem.
Para além disso são, se não o mais rápido, um dos animais terrestres mais velozes – atinge como pico de velocidade 112 km/h.
Esta especialização na velocidade acarretou perda de resistência e de robustez – as perseguições raramente duram mais de 15 segundos e escassas centenas de metros.
Socialmente, as chitas apresentam também algumas peculiaridades. Ao contrário de outros felídeos, em que o território ocupado pelos machos excede os das fêmeas, no caso das chitas as fêmeas são verdadeiras “rainhas” territoriais – no parque nacional do Serengueti verifica-se uma média de 833 km2 para as fêmeas contra apenas 37 km2 para os machos. As estratégias dos machos para a preservação de território passam por associação de vários indivíduos, geralmente grupos de irmãos.
Os imensos territórios ocupados pelas fêmeas, quando comparados com os dos machos, parecem facilitar o encontro de parceiros masculinos.
Para além de terem menos território, os machos apenas contactam com as fêmeas durante a época de acasalamento não contribuindo para a alimentação e protecção das crias.
 A análise genética das fezes das crias de chita, no Serengueti, permitiu concluir que em cada ninhada a paternidade é múltipla, ou seja, as fêmeas tinham copulado com vários machos.
A análise genética das fezes das crias de chita, no Serengueti, permitiu concluir que em cada ninhada a paternidade é múltipla, ou seja, as fêmeas tinham copulado com vários machos.
Este comportamento, poliandria, pode ser justificado para evitar a morte das crias por parte dos machos que não sejam os progenitores.
Por outro lado aumenta a variabilidade genética, potencialmente favorecedora de vantagens evolutivas na descendência.
A poliandria, parece ajudar igualmente a manutenção das coligações de machos pois, mantendo-se em grupos, terão maiores oportunidades de acasalamento.
O acasalamento das chitas é um fenómeno que raramente foi observado na natureza sendo estas conclusões obtidas indirectamente pela análise do património genético das crias.
Fonte: Gottelli D, Wang J, Bashir S & Durant SM (2007) Genetic analysis reveals promiscuity among female cheetahs. Proceedings of the Royal Society of London B doi:10.1098/rspb.2007.0502
Imagens: http://planet-earth.nnm.ru/dikie_koshki_i_kotyata_chast_5_gepard_remake
PATAS E GENES


Este estudo vem evidenciar que os genes envolvidos no desenvolvimento e aparição evolutiva dos autópodes (partes do esqueleto os carpos, metacarpos, metatarsos, metacarpos e falanges, resumindo, o ossos das “mãos” e “pés” de mamíferos, aves, répteis e anfíbios – os tetrápodes) já se encontravam no património genético há mais tempo do que quando se deu a transição para o ambiente terrestre..
Os estudos paleontológicos em exemplares de transição morfológica entre peixes, anfíbios e animais com verdadeiros membros locomotores deixavam em aberto a possibilidade daquela “revolução” evolutiva se ter dado de uma forma rápida em termos de tempo geológico – ver estudos em Acanthostega, Tiktaalik e Ichthyostega.
Este estudo molecular dos genes Hox (genes fundamentais reguladores do desenvolvimento em diferentes organismos e áreas do corpo, nomeadamente no desenvolvimento do esqueleto apendicular, ie, dos membros) vem mostrar que a “maquinaria” molecular necessária já estava presente. O que faltaria seriam apenas as condições ecológicas necessárias à sua verdadeira expressão.
Voltarei a este assunto mais tarde…
An autopodial-like pattern of Hox expression in the fins of a basal actinopterygian fish
Marcus C. Davis1, Randall D. Dahn1 & Neil H. Shubin1,2 Nature 447, 473-476 (24 May 2007)
Comparative analyses of Hox gene expression and regulation in teleost fish and tetrapods support the long-entrenched notion that the distal region of tetrapod limbs, containing the wrist, ankle and digits, is an evolutionary novelty. Data from fossils support the notion that the unique features of tetrapod limbs were assembled over evolutionary time in the paired fins of fish.
The challenge in linking developmental and palaeontological approaches has been that developmental data for fins and limbs compare only highly derived teleosts and tetrapods; what is lacking are data from extant taxa that retain greater portions of the fin skeletal morphology considered primitive to all bony fish. Here, we report on the expression and function of genes implicated in the origin of the autopod in a basal actinopterygian, Polyodon spathula. Polyodon exhibits a late-phase, inverted collinear expression of 5′ HoxD genes, a pattern of expression long considered a developmental hallmark of the autopod and shown in tetrapods to be controlled by a ‘digit enhancer’ region. These data show that aspects of the development of the autopod are primitive to tetrapods and that the origin of digits entailed the redeployment of ancient patterns of gene activity.
(Figuras : Heredity (2006) 97, 235-243. doi:10.1038/sj.hdy.6800872; published online 26 July 2006 Building divergent body plans with similar genetic pathways B J Swalla; http://en.wikipedia.org/wiki/Image:Fishapods.jpg)
Fósseis – História e mitos populares

Os fósseis ao longo dos tempos nem sempre foram encarados como registo de uma vida passada que permitem reconstituir tudo aquilo que se passou biologicamente no nosso planeta. Associações dos fósseis a acontecimentos históricos bem como tradições e mitos populares de várias partes do mundo são inúmeros. Alguns deles são aqui referidos.
Fóssil deriva do termo fossilis referido pela primeira vez por Plínio, o Velho (23-79 DC). A sua raiz fossus, particípio passado de fodere (i.e. cavar), significa literalmente “o que se extrai cavando“.
Adrienne Mayor refere no seu livro “The first fossil hunters” que na origem da figura mitológica Grifo estarão estado os dinossáurios. Senão vejamos o seu raciocínio: no séc. VII A.C., os gregos estabelecem contactos com nómadas Saka (exploradores de ouro no deserto de Gobi). Estes povos da Ásia central referiam que existia um monstro protector das reservas de ouro que teria cabeça e asas de águia num corpo de leão – é o nascimento da lenda do grifo na cultura grega. Nos anos 20 do séc. XX são descobertos dinossáurios no deserto de Gobi, um dos quais o Protoceratops – dinossáurio com uma projecção craneal semelhante a um bico.
A enorme semelhança de aspecto entre os restos de Protoceratops e a figura mitológica do grifo poderá explicar que os primeiros gregos (desconhecedores dos dinossáurios) tenham tomado conhecimento do seres do mesozóico muito antes de Richard Owen os definir no séc. XIX.
Outro dos exemplos históricos em que o registo fóssil e a história se cruzam diz respeito a Santo Agostinho (Aurelius Augustinus, 354-430 DC).
Em 413, no seu livro A Cidade de Deus, é referido um molar gigantesco atribuído a um gigante ancestral. Como outros autores até aí, pensava que os fósseis eram o resultado do Dilúvio. Acreditava igualmente que os seres humanos haviam diminuído de tamanho ao longo dos tempos. Esse molar seria um vestígio desses tempos em que os humanos apresentavam um tamanho colossal – hoje sabemos que esse molar não é mais do que o resto fossilizado de um parente dos actuais elefantes.
")
Belemnite
Em relação a um dos fósseis comuns no registo paleontológico português – belemnites – também existem várias crenças populares. As belemnites são o resto fossilizado de seres marinhos semelhantes a lulas e que habitavam o planeta nos tempos dos dinossáurios. Os restos que são preservados apresentam uma forma cónica, parecida com balas. É a sua forma que contribui para que vários povos expliquem a sua origem de maneiras distintas da real – resto de um ser vivo.
A designação inglesa para belemnite é thunderstone (pedra-de-raio) pois pensava-se que resultavam da queda de um relâmpago. No folclore chinês as belemnites são conhecidas como Jien-shih ou pedras-espada. Na Escandinávia aqueles fósseis são vistos como velas de elfos, gnomos ou de fadas. Nalgumas áreas ainda são actualmente designadas de vateljus que em sueco significa literalmente luzes de gnomo.
Outro modo de explicar o aparecimento de fósseis é a sua atribuição a fenómenos religiosos.
As amonites – moluscos cefalópodes marinhos semelhantes aos Nautilóides, existentes em várias afloramentos do país, exs: Figueira da Foz, Peniche.Na zona de Whitby, Inglaterra, considerava-se que eram restos petrificados de cobras que outrora haviam invadido esta área. A praga havia sido terminada por Santa Hilda (614-680 DC), que as transformou em rochas.
")
Orthosphynctes sp., Portugal
Associadas ao deus egípcio Ammon (representado por vezes com cornos retorcidos, de onde deriva o nome amonites), eram encaradas pelos gregos clássicos como símbolos sagrados capazes de curar mordeduras de cobra, cegueira, esterilidade ou impotência. Alguns romanos acreditavam que podiam prever o futuro se dormissem com uma amonite piritizada sob o travesseiro.
Dentes de tubarão fossilizados (geralmente de Carcharodon) eram utilizados como amuletos contra venenos. As designações tradicionais para estes fósseis incluíam Glossopetrae (língua de pedra), Linguae Melitensis (línguas de Malta) ou Linguae S. Pauli (línguas de São Paulo). Esta última é explicada pela seguinte associação entre um facto bíblico e a consequente exploração popular. Como referido em Actos dos Apóstolos (28:2-7), São Paulo, em Malta, foi mordido por uma cobra. Este atirou-a para a fogueira não tendo sofrido qualquer dano físico.
Como castigo divino as cobras terão perdido o seu veneno bem como os olhos e língua ficando para sempre os vestígios preservados sob a forma petrificada.
(Publicado no jornal O Primeiro de Janeiro a 29/09/2005)
Imagens: da Wikipedia, páginas de Belemnites e Amonites.


{kind=link}