


Cidades sem pessoas – I
 (Publicado no jornal O Primeiro de Janeiro a 02/08/2007 – continua amanhã)
(Publicado no jornal O Primeiro de Janeiro a 02/08/2007 – continua amanhã)
Cidade sem Sono
Ninguém dorme no céu. Ninguém, ninguém.
Não dorme ninguém.
As criaturas da lua cheiram e rondam as choupanas.
Virão iguanas vivas morder os homens que não sonham
e o que foge com o coração partido encontrará pelas esquinas
o incrível crocodilo imóvel sob o frouxo protestos dos astros.
(…)
Um dia
os cavalos viverão nas tabernas
e as formigas furiosas
atacarão os céus amarelos que se refugiam nos olhos das vacas.
Outro dia
veremos a ressurreição de mariposas dissecadas
e ainda, ao andar por uma paisagem de esponjas pardas e barcos mudos
veremos brilhar nosso anel e brotar rosas de nossa língua.
Alerta! Alerta! Alerta!
Aos que guardam ainda pegadas de garra e aguaceiro,
àquele rapaz que chora porque não sabe a invenção da ponte
ou àquele morto que já não tem mais que a cabeça e um sapato,
há que levá-los ao muro onde iguanas e serpentes esperam,
onde espera a mão mumificada do menino
e a pele do camelo se eriça com um violento calafrio azul.
(…)
Não dorme ninguém pelo mundo. Ninguém, ninguém.
Já o disse.
Não dorme ninguém.
Mas se alguém tem de noite demasiado musgo nas têmporas,
abri os alçapões para ver sob a lua
as falsas taças, o veneno e a caveira dos teatros.
Federico García Lorca
Este poema e um artigo no último número da Scientific American fizeram-me imaginar as cidades portuguesas sem os representantes da espécie humana.
De um momento para o outro e por qualquer motivo desconhecido, todas as pessoas desaparecem das cidades; surgirão vencedores e vencidos biológicos, no contexto citadino definitivamente abandonado pelos seus criadores.
No momento em que pela primeira vez a população urbana portuguesa ultrapassou a população rural, a especulação sobre o que aconteceria num ambiente urbano sem portugueses é curiosa, apresentando-se como uma realidade que deveremos tomar em atenção.
No I Congresso Europeu de Conservação Biológica, em 2006, foi discutido que “com metade da população mundial a viver actualmente em cidades e a previsão de 60% em 2030, o ambiente urbano é um dos pontos principais da agenda global de ambiente e de conservação.”
Mas o que se passaria nas nossas cidades, após o desaparecimento de todos os portugueses?
Os vencedores
Entre os vencedores biológicos deste omnicídio estão os mosquitos, que sem campanhas de extermínio, e aproveitando-se de zonas húmidas, aumentarão exponencialmente o seu número. Esses insectos alimentar-se-ão de animais como as aves. Estas, sem cabos de alta-tensão e arranha-céus (entretanto destruídos, por falta de manutenção), poderão voar livremente.
 Todos os anos os arranha-céus são responsáveis pela morte de 1000 milhões de aves, só nos EUA. Para Daniel Klem Jr., ornitólogo do Muhlenberg College, o revestimento em vidro dos edifícios das cidades constitui um fenómeno “indiscriminado, eliminando aptos e não-aptos”. As aves colidem com aquelas estruturas, pois não as conseguem identificar, por serem espelhadas, vendo apenas o céu ou árvores reflectidas. Segundo este investigador, só a destruição de habitats tem um impacto mais negativo sobre as aves.
Todos os anos os arranha-céus são responsáveis pela morte de 1000 milhões de aves, só nos EUA. Para Daniel Klem Jr., ornitólogo do Muhlenberg College, o revestimento em vidro dos edifícios das cidades constitui um fenómeno “indiscriminado, eliminando aptos e não-aptos”. As aves colidem com aquelas estruturas, pois não as conseguem identificar, por serem espelhadas, vendo apenas o céu ou árvores reflectidas. Segundo este investigador, só a destruição de habitats tem um impacto mais negativo sobre as aves.
E os nossos animais de estimação?
Os gatos contam-se entre os prováveis vencedores do período pós-humano, caçando pequenos mamíferos, insectos e aves, à semelhança do que fazem actualmente. Gradualmente aumentarão de tamanho, competindo directamente com outros predadores.
Os cães poderão ter dois destinos. As raças de comportamento dominante agrupar-se-ão em matilhas, à semelhança do que ocorre hoje em dia, em matilhas de cães assilvestrados. Desta forma poderão sobreviver, readquirindo alguns comportamentos ancestrais dos lobos, percorrendo ruas e avenidas em busca de presas. Presentemente, os cães abandonados são responsáveis por ataques a rebanhos que ocorrem no nosso país, embora os proprietários prefiram responsabilizar o lobo-ibérico…
Na minha opinião, os cães mais dóceis ou pequenos serão remetidos para nichos ecológicos reduzidos, ou terão como destino a extinção. O meu Labrador provavelmente não se safaria…
 Flora exótica como espanta-lobos (Ailanthus altíssima), robínia (Robinia pseudoacacia), acácia-de-espigas (Acacia longifolia) e árvore-do-incenso (Pittosporum ondulatum) existem nas nossas cidades, tendo sido aí introduzidas para ornamentação de jardins, arborização de espaços urbanos e sebes.
Flora exótica como espanta-lobos (Ailanthus altíssima), robínia (Robinia pseudoacacia), acácia-de-espigas (Acacia longifolia) e árvore-do-incenso (Pittosporum ondulatum) existem nas nossas cidades, tendo sido aí introduzidas para ornamentação de jardins, arborização de espaços urbanos e sebes.
A espanta-lobos é proveniente da China, produzindo até 350 000 sementes anualmente. Além de extremamente agressiva para as plantas autóctones, liberta toxinas que impedem o desenvolvimento de vegetação em seu redor.
Sem campanhas de erradicação, algumas plantas exóticas substituirão definitivamente as plantas originais, colonizando cada vez maiores áreas e modificando a paisagem natural.
Segundo Elizabete Marchante, do Centro de Ecologia Funcional da Universidade de Coimbra, “desde há pouco tempo, começou a surgir em espaços urbanos outra espécie (Sesbania punicea) que, apesar de ainda não ser invasora em Portugal, é uma invasora perigosa em ecossistemas com clima idêntico.” As árvores vencedoras do abandono humano ocuparão a maioria das ruas ao fim de dois a quatro anos. As suas raízes irão destruir progressivamente asfalto e passeios, bem como a rede de água e esgotos, contribuindo para o cada vez maior esquecimen
As árvores vencedoras do abandono humano ocuparão a maioria das ruas ao fim de dois a quatro anos. As suas raízes irão destruir progressivamente asfalto e passeios, bem como a rede de água e esgotos, contribuindo para o cada vez maior esquecimen
to dos vestígios humanos.
Nada do aspecto actual das cidades será mantido. Se houvesse alguém para o descrever, veria um ambiente caótico em que as marcas de construção humana seriam, pouco a pouco, engolidas pela vegetação.
(continua amanhã)
O mar e as entranhas – histórias de bactérias
 (Publicado no jornal O Primeiro de Janeiro a 12/07/2007)
(Publicado no jornal O Primeiro de Janeiro a 12/07/2007)
A escuridão é total. Mas há vida.
Não poderíamos viver nestes ambientes. Mas conhecemo-los.
Ou partes dele.
O fundo do mar.
O nosso sistema digestivo.
O grande desconhecido que é o profundo marinho tem equivalências no invisível interno humano.
O PNAS*, de 5 de Julho de 2007, publicou um estudo sobre a vida microscópica, onde se relacionam os dois ambientes, com personagens que estão mais relacionadas do que até aqui se imaginava.
Quer o sistema digestivo quer o fundo do mar são ambientes inóspitos – escuros e com baixas concentrações de oxigénio.
Ainda assim estão repletos de bactérias.
 Sulfurovum litthotrophicum, descrita em 1984, e Nitratiruptor tergarcus são duas espécies de bactérias do grupo ε-Proteobacteria, que habitam o fundo do mar. Sobrevivem a temperaturas comos as que temos no frigorífico lá de casa, 4º C, até aos 70º C. Vivem ambas no substrato marinho de grandes profundidades obtendo energia através da fixação de azoto provenientes de fontes hidrotermais. São consideradas das mais resistentes formas de vida, pois conseguem sobreviver naqueles ambientes adversos, onde as temperaturas podem atingir mais de 100º C, a profundidades, como no caso da fonte hidrotermal “Menez Gwen” dos Açores, de 1700 m.
Sulfurovum litthotrophicum, descrita em 1984, e Nitratiruptor tergarcus são duas espécies de bactérias do grupo ε-Proteobacteria, que habitam o fundo do mar. Sobrevivem a temperaturas comos as que temos no frigorífico lá de casa, 4º C, até aos 70º C. Vivem ambas no substrato marinho de grandes profundidades obtendo energia através da fixação de azoto provenientes de fontes hidrotermais. São consideradas das mais resistentes formas de vida, pois conseguem sobreviver naqueles ambientes adversos, onde as temperaturas podem atingir mais de 100º C, a profundidades, como no caso da fonte hidrotermal “Menez Gwen” dos Açores, de 1700 m.
Apesar da enorme resistência daquelas bactérias em ambiente natural, só recentemente foram cultivadas em laboratório permitindo que fossem estudadas mais detalhadamente.
O outro grupo de bactérias que ninguém gostaria conhecer, pelo menos na prática ocupa, com maior ou menor frequência, o nosso sistema digestivo.
 A Helicobacter pylori, descoberta em 1982, está presente em metade da população mundial, sendo a causadora da inflamação da mucosa do estômago bem como das úlceras gástricas e do duodeno. A descoberta desta relação concedeu, em 2005, o prémio Nobel da Medicina a Barry Marshall e J. Robin Warren.
A Helicobacter pylori, descoberta em 1982, está presente em metade da população mundial, sendo a causadora da inflamação da mucosa do estômago bem como das úlceras gástricas e do duodeno. A descoberta desta relação concedeu, em 2005, o prémio Nobel da Medicina a Barry Marshall e J. Robin Warren.
O minúsculo ser vivo acompanha a espécie humana desde há muito num fenómeno coevolutivo, facilitado pela sua grande variabilidade genética.
Segundo investigadores do Instituto Max Planck em Berlim, a Helibobacter tem sido transmitido de pais para filhos desde a nossa ancestral saída de África.
Reconstruindo a árvore evolutiva desta bactéria, foi possível identificar dois grandes ramos – um que infecta os europeus e norte-americanos e outro que afecta sobretudo os asiáticos. Essas duas linhagens estão associadas às migrações humanas, permitindo reconstituir essas antigas movimentações.
 Outro dos géneros de bactérias patogénicas analisado foi o Campylobacter jejuni, responsável por intoxicações alimentares nomeadamente a gastroenterite. Os ambientes favoritos para a sua disseminação são leite cru ou mal pasteurizado, aves mal cozinhadas e água não tratada (líquida ou em gelo).
Outro dos géneros de bactérias patogénicas analisado foi o Campylobacter jejuni, responsável por intoxicações alimentares nomeadamente a gastroenterite. Os ambientes favoritos para a sua disseminação são leite cru ou mal pasteurizado, aves mal cozinhadas e água não tratada (líquida ou em gelo).
A equipa de investigadores procedeu à análise do ADN presente nas bactérias que partilham o nosso ambiente digestivo – Helicobacter e Campylobacter – e o das bactérias das profundezas marinhas – Sulfurovum e Nitratiruptor.
Os dois grupos de bactérias apresentaram afinidades genéticas, que lhes possibilitam viverem em ambientes hostis. Entre as semelhanças estão a quase ausência de genes de reparação do ADN. Este facto permite não só a grande adaptação destes seres vivos a novas condições extremas, mas também ao próprio sistema de defesa de um organismo hospedeiro.
 Segundo os investigadores, as bactérias humanas evoluíram a partir de ancestrais de grande profundidade, adquirindo o seu “mau-feitio” quando estabeleceram relações simbióticas com invertebrados marinhos.
Segundo os investigadores, as bactérias humanas evoluíram a partir de ancestrais de grande profundidade, adquirindo o seu “mau-feitio” quando estabeleceram relações simbióticas com invertebrados marinhos.
Não era novidade que tínhamos todos uma origem marinha.
Foi de lá que viemos.
O profundo marinho e o sistema digestivo humano partilham coincidências evolutivas.
O sistema digestivo humano serve de mar a uma variedade de fauna microscópica; sabemos agora que parte dessa fauna tem parentes próximos nos fundos marinhos.
* Proceedings of the National Academy of Sciences
Referências consultadas
-Inagaki, F. et al. 2004. Sulfurovum lithotrophicum gen. nov., sp. nov., a novel sulfur-oxidizing chemolithoautotroph within the ε -Proteobacteria isolated from Okinawa Trough hydrothermal sediments. International Journal of Systematic and Evolutionary Microbiology, 54, 1477-1482.
-Nakagawa, S. et al. 2007. Deep-sea vent ε-proteobacterial genomes provide insights into emergence of pathogens PNAS published July 5, 2007, 10.1073/pnas.0700687104.
-Nakagawa, S. et al. 2005. Nitratiruptor tergarcus gen. nov., sp. nov. and Nitratifractor salsuginis gen. nov., sp. nov., nitrate-reducing chemolithoautotrophs of the -Proteobacteria isolated from a deep-sea hydrothermal system in the Mid-Okinawa Trough. International Journal of Systematic and Evolutionary Microbiology, 55, 925
Auto da Ocorrência
 (Publicado no jornal O Primeiro de Janeiro a 28/06/2007)
(Publicado no jornal O Primeiro de Janeiro a 28/06/2007)
Trânsito parado. Avançamos. Polícias. Medem algo na estrada. Mais à frente, carros batidos. Os peritos continuam a medir e a escrever. Finalmente conseguimos passar, apenas retendo na memória o resultado final do que aconteceu.
A familiar cena de cidade poderia ser um qualquer dia de um paleontólogo que estude pegadas de dinossauro.
Tal como os polícias e os mirones, também os cientistas procuram saber o que se passou. Medem os vestígios do acontecimento ocorrido algures num passado mais ou menos remoto. Os elementos da autoridade medem o rasto da travagem para inferirem o tempo de duração da mesma e a velocidade provável a que se deslocava o carro. Os paleontólogos medem o espaço entre pegadas para deduzirem a velocidade do animal. Esta medição permitiu, por exemplo, constatar que, no Cabo Espichel, o trilho de um dinossauro carnívoro apresentava um passo (série de duas pegadas) irregular; por outras palavras, o animal coxeava. As razões para este comportamento podem ser várias: ferimento numa das patas ou, motivo mais difícil de comprovar, poderia estar a transportar uma presa.
Os paleontólogos conseguem inferir uma série de informações biológicas a partir da “cena do crime”: p.e., com base no tamanho e forma da pegada, conseguem concluir a altura do animal até à anca e assim ter uma ideia geral do tamanho do animal.
 No caso da maior jazida portuguesa de pegadas de dinossauro – Pedreira do Galinha, na zona de Fátima – podem observar-se centenas de pegadas de saurópodes – dinossauros herbívoros quadrúpedes. Este local apresenta os maiores rastos de dinossauros do Jurássico médio (sensivelmente há 165 milhões de anos) a nível mundial, dois dos quais com mais de 140 metros de extensão.
No caso da maior jazida portuguesa de pegadas de dinossauro – Pedreira do Galinha, na zona de Fátima – podem observar-se centenas de pegadas de saurópodes – dinossauros herbívoros quadrúpedes. Este local apresenta os maiores rastos de dinossauros do Jurássico médio (sensivelmente há 165 milhões de anos) a nível mundial, dois dos quais com mais de 140 metros de extensão.
Não só os rastos permitem deduzir informações sobre a velocidade e comportamento do animal. A partir da forma das pegadas individuais, os paleontólogos obtêm informações sobre o seu autor: à semelhança de um CSI natural, deduzem, com maior ou menor rigor, o retrato-robô de quem andou (literalmente) num determinado local.
As pegadas de dinossáurio também exerceram fascínio na produção literária.
 A descoberta em 1909 de pegadas de Iguanodon, em Inglaterra, originou uma enorme excitação em Sir Arthur Conan Doyle, o criador do detective Sherlock Holmes.
A descoberta em 1909 de pegadas de Iguanodon, em Inglaterra, originou uma enorme excitação em Sir Arthur Conan Doyle, o criador do detective Sherlock Holmes.
Alguns autores apontam este motivo, bem como a publicação da “Origem das Espécies” de Darwin, como os principais factores de inspiração para que Conan Doyle escrevesse “O Mundo Perdido”, relato de aventuras num país da América do Sul povoado de criaturas perigosas e pretensamente extintas.
Ao contrário do que se passa nos acidentes de automóvel, em que os responsáveis materiais normalmente ficam junto do local da “ocorrência”, no caso das pegadas de dinossauro estes nunca lá estão para soprar no balão. Uma das perguntas mais frequentes que me são feitas refere-se ao motivo pelo qual os ossos de dinossauro nunca são encontrados perto das jazidas de pegadas. As razões são essencialmente duas: a maioria das pegadas é produzida em momentos de actividade biológica habitual, isto é, quando o animal se encontrava em movimento para pastar ou caçar, não sendo provável, assim, que tivesse deixado aí o seu esqueleto…
O segundo motivo diz respeito às condições de preservação – tafonomia – dos vestígios. Pegadas e ossos necessitam de condições geológicas diferentes para fossilizar, ou seja, os ingredientes para a fossilização são distintos para o registo icnológico (pegadas) e o registo osteológico (ossos).
Tal como os índios norte-americanos, que perseguiam os seus adversários ou as presas numa caçada, também os paleontólogos seguem os rastos, embora nunca consigam alcançar os seus autores… ao contrário daqueles, apenas ficam com pedaços duma ocorrência do tempo passado.
Foto: Luís Azevedo Rodrigues – jazida de Vale de Meios
Baleias com patas, a Origem das Espécies e aves fósseis
 (Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 24/11/2005)
Hoje comemoram-se 146 anos de existência de um livro que teve fortes repercussões pelas suas propostas de Evolução pelo mecanismo de Selecção Natural. A 24 de Novembro de 1859, Charles Darwin publicou “A Origem das Espécies” com uma primeira edição de 1250 exemplares que esgotou no seu dia de lançamento.
Darwin defendia que o meio ambiente e as relações entre os próprios seres vivos exercem uma selecção que favorece os mais aptos enquanto os menos dotados são eliminados, transmitindo-se à geração seguinte as diferenças que facilitam aquela sobrevivência. Ao longo das gerações, essas características acentuam-se e geram uma nova espécie.
Darwin foi convidado em 1831 a participar numa volta ao mundo no navio Beagle promovida pela marinha inglesa. A expedição – que durou cinco anos – tinha o objectivo de melhorar e completar dados cartográficos. Esta viagem foi decisiva para fundamentar as suas teorias evolutivas.
América do Sul, Austrália e Nova Zelândia foram alguns dos locais em que Beagle efectuou paragens. Surpreendeu-se com o grande número de espécies de plantas e de animais que, até então, eram desconhecidos. O que lhe chamou mais atenção foram a enorme diversidade de tentilhões, que só conheceu no arquipélago das Galápagos.
 O registo fóssil
O registo fóssil
Quando Darwin propôs que a Evolução se dava pela Selecção Natural, o registo fóssil oferecia ainda poucas evidências que apoiassem as suas ideias.
Actualmente os paleontólogos contam com mais informação fossilífera do que aquela que dispunha Darwin. Este dedicou dois capítulos da sua “Origem das Espécies” à imperfeição do registo fóssil, provavelmente porque constatava que estaria aí um dos pontos mais fracos da sua argumentação. Dois anos depois da publicação do seu livro, o primeiro exemplar de Archaeopteryx foi descoberto na Baviera, constituindo um dos “elos perdidos” entre as aves e os répteis.
O registo fóssil está longe de ser perfeito – faltam assim muitos elementos de transição na história evolutiva dos seres vivos. Este é um dos argumentos com que os Criacionistas (opositores à Teoria da Evolução, populares sobretudo nos EUA) se valem no seu ataque a Darwin.
As últimas estimativas apontam para que apenas 1% de todas as espécies animais e vegetais que habitaram o nosso planeta ficaram preservadas como fósseis. Razões para esta escassez de informação fóssil são várias, mas podemos apontar que as condições físicas necessárias à preservação de um ser vivo sob a forma de fóssil são muito raras. Grande parte dos seres vivos é constituída por partes moles; este facto contribui igualmente para que o registo fóssil seja desprovido da maioria dos “actores” da vida sobre a Terra.
Mas existem numerosos exemplos que reflectem as etapas de transição entre diversos grupos de organismos, corroborando Darwin.
Para além do já referido Archaeopteryx (descoberto em sedimentos da Baviera com 150 milhões de anos) foi descoberta nos últimos 20 anos toda uma panóplia de formas de transição entre os dinossáurios carnívoros (semelhantes ao Velociraptor) e as actuais aves: Confuciusornis (China, primeira evidência de um bico sem dentes); Iberomesornis (Espanha, apresentava uma estrutura óssea semelhante às aves actuais que permitia inserir a musculatura específica para o voo), entre inúmeros outros exemplos.
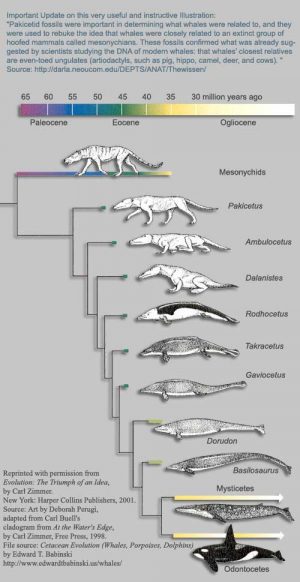 As patas das Baleias
As patas das Baleias
Outro dos exemplos que se podem apresentar para ilustrar as transições evolutivas sofridas pelos organismos é a evolução das baleias.
A maioria das pessoas tem, pelo menos a noção, de que a vida terrestre surgiu a partir de vertebrados que deixaram o ambiente aquático. Todas as formas de animais que ocuparam e ocupam um ambiente terrestre, descendem desses primeiros colonizadores. O que a maioria das pessoas não sabe é que o grupo de animais a que pertencem as actuais baleias descendem de um grupo que “decidiu” voltar a ambientes aquáticos, donde tinham saído há mais 250 milhões de anos.
Em 1983 foram descobertos no Paquistão fósseis de um animal que tinha vivido há cerca de 52 milhões de anos. Este animal, Pakicetus, apresentava ainda corpo com forma para a vida terrestre (membros com capacidade de locomoção em terra) mas tinha um crâneo e dentes com características típicas dos ancestrais dos actuais baleias. Onze anos mais tarde e igualmente no Paquistão foi descoberto o Ambulocetus natans (literalmente baleia caminhante que nada). O Ambulocetus tinha o tamanho de um leão-marinho e apresentava as patas (sim esta baleia ancestral tinha patas!) com capacidades para deslocação em ambiente terrestre. Igualmente exibia os seus pés e mãos com capacidade natatória – ou seja este animal possuía capacidades para se deslocar em ambiente terrestre e aquático.
Aparentemente o Ambulocetus nadava como uma lontra, com movimentos para cima e para baixo (dados da morfologia da sua coluna vertebral atestam-no).
Em 1995 um terceiro elemento de transição foi descoberto – o Dalanistes. Apresentava os membros mais curtos que Ambulocetus, cauda e crâneo mais alongados ou seja mais semelhantes às actuais baleias.
Actualmente mais de uma dúzia de fósseis ilustrativos das transições evolutivas dos cetáceos (grupo a que pertencem as baleias) já foram descobertos.
Complementarmente análises de ADN mitocondrial aos actuais representantes dos cetáceos permitem apontar que estes pertencem ao grupo dos artiodáctilos, mais concretamente são parentes próximos dos hipopótamos.
No dia de aniversário da A Origem das Espécies, que fez com que o Homem descesse de mais um dos seus inúmeros pedestais, podemos afirmar que o registo fóssil é mais um dos motivos de orgulho para Darwin.
Ao contrário do que receava, o trabalho de investigação paleontológico nos últimos 100 anos, permitiu que o registo fóssil seja mais uma prova de que Darwin não estava e não está errado.
Imagem – Werner Horvath: “Charles Robert Darwin – Evolution”. Oil on canvas
A Evolução escondida nos Cartoons
 (Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
(Publicado no jornal O Primeiro de Janeiro a 10/11/2005)
A História da vida animal está cheia de alterações na forma dos seus corpos. A análise da forma (análise morfológica) é um das ferramentas no estudo da história evolutiva dos seres vivos.
Um dos essenciais evolucionistas do nosso século – Stephen Jay Gould da Universidade de Harvard – utilizou o melhor da dedução científica para analisar um dos ícones da Disney.
Gould recolheu uma sequência cronológica de Ratos Mickey, desde os anos 20 até à actualidade, analisando alguns traços morfológicos desta figura da banda-desenhada – altura e tamanho geral do crâneo, bem como o tamanho dos olhos. Constatou que ao logo das mais de cinco décadas da história do pequeno ratinho, os parâmetros analisados tinham aumentado. Por outras palavras, Mickey tinha-se transformado: de uma figura de crâneo pequeno e alongado e de olhos pequenos num ratinho de crâneo arredondado e alto com olhos proporcionalmente grandes. Se olharmos para a figura qual dos dois extremos “evolutivos” de Mickey nos parece mais atractivo?
A maioria das pessoas referirá que o da direita (olhos maiores, crâneo mais arredondado). Estas características morfológicas podem ser reconhecidas, na grande maioria dos casos, como sendo características de crias de mamíferos. Todos nós facilmente identificamos que os bebés apresentam um crâneo e olhos proporcionalmente maiores do que os adultos. Estas particularidades têm um efeito sobre os membros de uma determinada espécie e igualmente nos humanos. Características morfológicas de juvenis parecem desencadear reacções de protecção e carinho – um crâneo grande e arredondado, olhos enormes, mandíbula pequena, etc.
Estes são alguns dos traços morfológicos apelativos nos mamíferos. Desta maneira Gould constatou que os desenhadores da Disney alteraram a fisionomia de Mickey atribuindo-lhe características morfológicas juvenis com o objectivo o tornarem emocionalmente mais apelativo.
Pelo contrário, podem ser encontradas algumas morfologias opostas em alguns dos vilões da Disney. Estes apresentam características anatómicas geradoras de desconfiança – crâneo afilado, olhos pequenos, mandíbula proeminente. Enquadram-se neste grupo a Rainha da Branca de Neve, a bruxa da Bela Adormecida e o feiticeiro Jafar em Aladino.
Mas o que tem isto a ver com a evolução?
Gould não perdeu o seu tempo com esta actividade aparentemente estéril. Pretendia ilustrar, com exemplos facilmente reconhecíveis, alguns conceitos da morfologia evolutiva. Uma das noções subjacentes ao estudo dos Mickeys é o da Pedomorfose – aquisição no adulto de um grupo descendente de características morfológicas juvenis do grupo antepassado.
 Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Existem alguns exemplos muito evidentes de Pedomorfose – a salamandra mexicana Ambystoma mexicanum exibe quando adulta guelras, prova morfológica juvenil dos seus ancestrais.
Este conceito está englobado num conjunto maior que é a Heterocronia – em sentido geral, a variação no tempo de aparecimento de uma característica morfológica de um grupo descendente comparativamente ao aparecimento dessa característica no grupo antepassado.
De uma maneira mais simples: podemos ver a “quantidade de transformação” que um indivíduo sofre ao longo do seu desenvolvimento é a mesma, maior ou menor do que do seu antepassado (aqui em sentido de evolutivo e não de progenitor).
O estudo da Heterocronia tem chamado a atenção nos últimos anos dos investigadores da Evolução. É um campo complexo mas muito apelativo e em que têm sido feitas inúmeras descobertas no estudo da morfologia de seres vivos actuais ou dos seus antepassados fósseis.
Outro dos exemplos de Pedomorfose no mundo não-natural é a evolução do símbolo da Shell. O seu logotipo é o bivalve Pecten tendo este “evoluído” desde o início do século por redução do número de “linhas” na concha, aparecendo assim menos complexo e mais juvenil.
Mas existem muito exemplos que podem ilustrar os conceitos evolutivos já referidos: é fazer uma busca na Internet pela “evolução” do Snoopy…

Patas para que vos quero!

Mas e se fosse um peixe?
“Dois belos pés!” afirma o comentador desportivo, numa tarde de futebol.
Nós temos. Os peixes não.
A “simples” diferença na forma do esqueleto, como ter “mãos” e “pés” ou autópodes, carrega uma importante história evolutiva desde os peixes até aos animais como nós.
Ao segurar um jornal, o leitor está, em termos evolutivos, a utilizar uma barbatana muito complexa e evoluída, e pertence a um grupo de vertebrados chamados tetrápodes, animais com quatro membros, que incluem animais como os mamíferos, aves, répteis e anfíbios.
O aparecimento dos ossos dos dedos em alguns anfíbios deveria ser resultado de nova “maquinaria” genética, pois todas as estruturas orgânicas são o resultado da informação que está contida nos genes. Será assim?
Um estudo publicado, a 24 de Maio, na revista Nature, refere que os genes necessários à formação dos dedos das “mãos” e “pés” dos tetrápodes têm uma história que remonta há 360 milhões de anos ou seja antes de os animais terem feito a “invasão” da terra. O estudo molecular dos genes HoxD (genes reguladores do desenvolvimento em diferentes organismos e áreas do corpo, concretamente no desenvolvimento do esqueleto apendicular, i.e., dos membros) vem mostrar que o património genético necessário já estava presente em peixes primitivos como o actual peixe actinopterígeo Polyodon spathula, considerado um autêntico fóssil vivo.
A análise genética deste animal permitiu afinar as informações paleontológicas com as da biologia do desenvolvimento, possibilitando que estas analisassem dados genéticos de peixes menos “evoluídos” – os actinopterígeos – e os comparassem com os dos tetrápodes. Tradicionalmente, estas análises eram efectuadas em peixes mais “evoluídos”, os teleósteos.
Os estudos paleontológicos em exemplares de transição morfológica entre peixes e animais com verdadeiros membros locomotores deixavam em aberto a possibilidade daquela “revolução” evolutiva se ter dado de uma forma rápida em termos de tempo geológico mas o Polyodon revelou que o património genético que permitiu o aparecimento de verdadeiras patas é mais antigo do que se suponha.
Fundamental para se compreender esta “novela” científica é o conhecimento dos fósseis de transição deste trajecto evolutivo.
Os “fotogramas” que permitem visualizar as alterações morfológicas entre as barbatanas e verdadeiros membros locomotores são vários. Conhecia-se já há algum tempo a parte mais inicial do “filme” – os peixes Eusthenopteron e Panderichthys – e a mais avançada – os anfíbios do Devónico superior como Acanthostega e Ichthyostega. Recentemente foi descoberto mais um “fotograma” – o peixe Tiktaalik; este, apresenta um mosaico de características morfológicas antigas e modernas, no trajecto evolutivo para o aparecimento de verdadeiros autópodes.
Algumas curiosidades morfológicas destes “primos” afastados: Ichtyostega possuía sete dedos em cada pata; o Acanthostega, oito. Desculpem mas não resisto a dizer: “vão-se as barbatanas mas fiquem os dedos!”.
Da próxima vez que um qualquer criacionista falar em falta de fósseis de transição nada como descrever estes belos nomes – Eusthenopteron, Panderichthys, Acanthostega, Tiktaalik e Ichthyostega!
Um outro estudo, de 2006 e desta vez embriológico, levado a cabo em Barcelona, permitiu analisar o processo de formação e disposição de dois ossos do pé em embriões humanos – o calcâneo (osso que constitui o nosso calcanhar) e o astrágalo, ambos ossos do pé.
Foram descritas semelhanças morfológicas entre um embrião humano de 33 dias, nas extremidades inferiores, com barbatanas; aos 54 dias o calcâneo e o astrágalo estão localizados no mesmo preciso local que em Bauria cynops, um réptil mamaliforme que viveu há 260 milhões de anos. As semelhanças anatómicas de posicionamento às 8 semanas e meia dos ossos referidos são enormes entre o embrião humano e a espécie fóssil Diademodon, que viveu há 230 milhões de anos.
Os autores deste estudo afirmam, que nesta fase, o posicionamento, e consequências ao nível da locomoção, dos ossos analisados estão a meio “caminho” entre répteis e mamíferos. Este tipo de análises incrementa o conhecimento morfológico efectuado por vários autores no séc. XIX, mesmo antes de Darwin publicar a sua obra magna, como Karl Ernst von Baer (1792-1876), que notou semelhanças morfológicas entre embriões de grupos diferentes. Conta a “tradição”, que von Baer, trabalhava no seu gabinete, e encontrou dois frascos com embriões de aves e lagartos; sem rótulos, não os pôde distinguir à primeira vista…
 Von Baer propôs que estádios embrionário iniciais conservavam padrões morfológicos comuns a vária espécies sendo os estádios mais avançados reveladores de divergência morfológica – as similitudes observadas entre embriões humanos e espécies do passado comprovam que anda bem que os frascos de von Baer deveriam ter os rótulos! Resumindo: espécies que divergem morfologicamente em estádios mais iniciais irão ser morfologicamente mais distintas em estádios adultos.
Von Baer propôs que estádios embrionário iniciais conservavam padrões morfológicos comuns a vária espécies sendo os estádios mais avançados reveladores de divergência morfológica – as similitudes observadas entre embriões humanos e espécies do passado comprovam que anda bem que os frascos de von Baer deveriam ter os rótulos! Resumindo: espécies que divergem morfologicamente em estádios mais iniciais irão ser morfologicamente mais distintas em estádios adultos.Von Baer foi pioneiro nas propostas que fez ao nível do desenvolvimento embrionário sendo o seu trabalho basilar numa das áreas mais importantes das Biologia actual – a evolução e o desenvolvimento, Evo-Devo.
De tudo o que vimos só me resta afirmar que a as teorias evolutivas que explicam o nosso trajecto na história da Terra têm cada vez mais “pés para andar…!”
(Publicado no jornal O Primeiro de Janeiro a 31/5/2007)
BIBLIOGRAFIA
Carroll, R.L., Irwin J. & Green, D.M. 2005. Thermal physiology and the origin of terrestriality in vertebrates. Zool. J. Linn. Soc. 143: 345-358.
Carroll, S. B. 2005. Endless Forms Most Beautiful: The New Science of Evo Devo and the Making of the Animal Kingdom, W. W. Norton & Company.
Davis, M.C. et al. 2007. An autopodial-like pattern of Hox expression in the fins of a basal actinop
terygian fish. Nature 447: 473-476.
Evo-Devo – http://www.pnas.org/cgi/content/full/97/9/4424
Goodwin, B. 1994. How the Leopard Changed its Spots, Phoenix Giants.
Isidro, A. & Vazquez, M.T. 2006. Phylogenetic and ontogenetic parallelisms on talo-calcaneal superposition. The Foot 16, 1-15.
FIGURAS:
Carroll, R.L., Irwin J. & Green, D.M. 2005. Thermal physiology and the origin of terrestriality in vertebrates. Zool. J. Linn. Soc. 143: 345-358.
Clack, J. 2002. An early tetrapod from Romer’s Gap, Nature 418, 72 – 76.
Horder, T.J. 2006. Gavin Rylands de Beer: how embryology foreshadowed the dilemmas of the genome. Nat Rev Genet. 7(11):892-8.
Grandes Opções

(Publicado no jornal O Primeiro de Janeiro a 10/5/2007)
Situações há que impõem que se façam escolhas e renúncias.
Já abordei algumas dessas opções, num contexto biológico, no texto “Tempos de crise – apertar o coração e o fígado”.
Eram referidas modificações fisiológicas que uma gazela do Deserto da Arábia sofre para poder suportar as duras condições daquele meio.
Contudo, não são só as condições adversas do meio-ambiente que impõem mudanças na fisiologia do organismo.
Uma das funções essenciais para todos os organismos é a reprodução – transmitir os seus genes à descendência – numa perspectiva que Richard Dawkins apelidou de O Gene Egoísta, no livro homónimo.
O Antechinus stuartii é um pequeno marsupial carnívoro que apresenta, no seu comportamento reprodutivo, alterações fisiológicas que, na maioria dos casos, conduzem a um desfecho trágico.
Tal como outros marsupiais, o Antechinus é um animal solitário e nocturno, caçando sobretudo insectos, mas também pequenas aves e répteis.
 A época de reprodução deste mamífero inicia-se no final do Inverno australiano, numa época de escassez alimentar, surgindo então as primeiras modificações corporais. Um mês antes da época de acasalamento, o corpo do Antechinus macho suspende a produção de esperma. O aparente paradoxo desta mudança fisiológica pode ser explicado pela redução nos gastos energéticos. Aquele fenómeno exige do animal um dispêndio considerável de energia na produção do material biológico envolvido na reprodução.
A época de reprodução deste mamífero inicia-se no final do Inverno australiano, numa época de escassez alimentar, surgindo então as primeiras modificações corporais. Um mês antes da época de acasalamento, o corpo do Antechinus macho suspende a produção de esperma. O aparente paradoxo desta mudança fisiológica pode ser explicado pela redução nos gastos energéticos. Aquele fenómeno exige do animal um dispêndio considerável de energia na produção do material biológico envolvido na reprodução.
Mas as mudanças não ficam por aqui.
Para além de suspender a produção de esperma, este animal inibe igualmente a síntese de algumas proteínas fundamentais, reduzindo ainda a actividade do próprio sistema imunitário. Estes fenómenos visam igualmente minimizar o gasto energético, com vista ao acto reprodutivo.
No entanto, a alteração verdadeiramente surpreendente é o enorme aumento de concentração sanguínea de testosterona. Esta hormona, produzida nos testículos e glândulas supra-renais, é conhecida popularmente como a hormona masculina e está associada ao aumento da libido e da agressividade.
Resumindo: o Antechinus macho cessa parte das actividades metabólicas, com o objectivo de preservar a sua energia, ao mesmo tempo que aumenta a produção de hormonas ligadas à necessidade obsessiva de se reproduzir.
Apesar de solitário durante a maior parte do tempo, a época reprodutiva é bastante activa, chegando mesmo a ser violenta. Durante esse período, os machos socializam, formando “arenas de cortejamento” onde disputam as fêmeas.
Os machos procuram freneticamente as fêmeas, utilizando a denominada cópula prolongada – entre 5 e 14 horas – com o fim de evitar que outros machos copulem.
Contudo, todas aquelas alterações fisiológicas têm custos biológicos. Após a época de acasalamento, os machos apresentam-se doentes e com parasitas, devido à depressão a que havia sido sujeito o seu sistema imunitário, observando-se que a maioria não sobrevive ao primeiro ano. Pelo contrário, em cativeiro, os Antechinus podem atingir cinco anos de longevidade.
Há, assim, um investimento biológico quase total na reprodução, tão grande que os machos não sobrevivem ao primeiro ano e aqueles que sobrevivem apresentam-se estéreis e sem viabilidade.
Os “super-machos”, carregados de testosterona, ficam desta forma condenados pelas suas opções.
Tal como os Antechinus “cortam” parte das suas funções vitais para poderem ter prole, apesar das consequências trágicas, também uma sociedade que tem de fazer cortes enormes em sectores fundamentais como a Educação e a Saúde pode correr o risco de não sobreviver.
Desejo que essas opções conduzam não à morte, mas à sobrevivência da “espécie” Portugal.
Referências
Kerr, J.B., and Hedger, M.P. (1983). Spontaneous spermatogenic failure in the marsupial mouse Antechinus stuartii Macleay (Dasyuridae, Marsupialia). Austral. J. Zool. 31, 445-466.
Bradley, A. J., I. R. McDonald, et al. (1980). Stress and mortality in a small marsupial (Antechinus stuartii, Macleay). Gen Comp Endocrinol 40(2): 188-200.
Ilustrações
Rui Ricardo e ESD
Os Ossos do meu Ofício

Ao longo dos últimos anos parte do meu trabalho tem consistido em estudar as colecções de dinossáurios em diversos Museus de História Natural de vários países e continentes a fim de compreender a evolução daqueles animais.
Os objectos de estudo são assim ossos fossilizados que têm que ser observados, medidos, estudados, mexidos e remexidos.
Percorri uma parte substancial da China, desde Pequim às províncias de Chengdu e Yunnan. Neste última estive em Lufeng, uma pequeníssima cidade, para os standards chineses, com o objectivo de estudar os famosos exemplares do Triásico superior (há cerca de 200 milhões de anos).
As condições de trabalho não eram os habituais de forma que acabei digitalizar os exemplares em cima de uma mesa de pingue-pongue. Apesar do carácter pouco ortodoxo do equipamento, a estrutura funcionava tão bem como a melhor mesa de trabalho do Museu de História Natural de Nova Iorque!
Lá se foi a minha idealização da curiosidade científica da classe trabalhadora…
 No início deste ano estive a estudar e digitalizar as colecções de dinossáurios do Museu de História Natural de Londres (MHNL).
No início deste ano estive a estudar e digitalizar as colecções de dinossáurios do Museu de História Natural de Londres (MHNL).
Entre os exemplares estudados contavam-se alguns saurópodes procedentes das jazidas de Tendaguru, na actual Tanzânia. Já havia estudado parte destas colecções no Museu de História Natural de Berlim – o mais famoso representante daqueles dinossáurios é o Brachiosaurus, cujo úmero (osso que vai do obro ao cotovelo) tem mais de dois metros!
Foram os alemães nos anos 20 do séc. XX, em especial expedições lideradas por Janensch, os principais exploradores das jazidas da Tanzânia. Mas igualmente os ingleses, sensivelmente na mesma época, fizeram expedições paleontológicas naquela área.
Encontrava-me a preparar o digitalizador 3D para enfrentar um fémur de um Dicraeosaurus, na cave do Museu, quando entra a responsável (curadora) pelas colecções de répteis do museu londrino.
Para além do rotineiro discurso sobre as localizações e características do material diz-me:
“Os ossos para além de muito grandes são também radioactivos!”
Reparo agora que tinha entrado pelas colecções de contador Geiger na mão!
Esperava ver quase tudo menos um contador de radioactividade.
“Agora põe estas luvinhas para manusear o material.”
Já estava eu agarrado literalmente ao osso quando a curadora me repete “Tem que pôr as luvas!”
Calma, disse de mim para mim.
“Madam, eu fumo mais de dois maços de tabaco, não sou abstémico nem vegetariano, estive dois meses na China e em Berlim trabalhei com o mesmo tipo de ossos, vivo em Portugal e respectiva crise económica…acha que é este mísero osso em que vou trabalhar na próxima meia-hora que me vai matar?!!?”
Senti uma pequena alteração comportamental na sua habitual calma, mas nada de especial. Que nos outros museus ela não tem nada com isso mas que ali são as normas, disse-me ela.
“Então já que tenho que usar as luvas ao menos quero ver esse brinquedo do meu imaginário funcionar! Pode ser?!”
Suspiro (dela, claro).
Baixa-se, começa a medir o fémur, e tudo abaixo do nível recomendado pelos britânicos (já de si ridiculamente baixos, citando um paleontólogo amigo, “tem mais radioactividade um pacote de amendoins…”).
“Muito bem…então agora meça-me a mim!”, disse-lhe, do alto da minha chica-espertice.
A fleuma britânica viu-se neste momento pois pressenti que por dentro do seu ar esfíngico pensava:
“Mas porque raio o comité científico atribuiu uma bolsa a este tipo?”
Começa a medir-me e, das duas uma, ou estou carregadinho de radiação ou os ossos são tão radioactivos como um esquilo siberiano a comer um chupa-chups!
“Minha senhora, o meu caso está encerrado. Posso trabalhar e tirar as luvas?”
E assim fiz.
Obviamente que não critico, antes pelo contrário apoio, as medidas de segurança adoptadas no NHML.
Neste momento penso que não deveria ter tido aquela atitude mas por vezes acho que os meus ossos são mais fortes que o meu ofício.
Espero.
(Publicado no jornal O Primeiro de Janeiro a 26/4/2007)
Imagens: Luís Azevedo Rodrigues
Ir

O trabalho científico, para o público em geral, é feito no recato de laboratórios e gabinetes. Um trabalho rotineiro, de pequenas ou grandes práticas, em ambientes idênticos, dia após dia.
No caso da Paleontologia de Vertebrados esses procedimentos podem ser iguais ao de qualquer investigador ou mesmo de qualquer profissional.
Mas também podem ser totalmente distintos.
Uma das componentes de um paleontólogo de vertebrados (mas não só) envolve a recolha de amostras dos animais que estudamos (ossos, na maioria das vezes, mas também dentes, pele, são analisadas pegadas, entre outros vestígios fossilizados).
A prospecção e recolha dos fósseis implica que o paleontólogo se tenha que deslocar às jazidas rochosas onde previamente já foram descobertos vestígios ou novas jazidas que, pelas características rochosas (litologia, idade, etc.), apresentem boas possibilidades de se mostrarem produtivas.
Em termos práticos os paleontólogos têm que ir para o campo!
Esse é uma dos elementos que a maioria dos paleontólogos mais aprecia e anima.
Para além do potencial profissional que pode implicar (novas espécies ou melhor e maior quantidade de material fossilizado) existe um lado inerente à sua actividade, e partilhado por outros cientistas das Ciências Naturais, que os enriquecem como pessoas.
Falo do contacto implícito com a Natureza.
Apesar de todas a contrariedades inerentes – por vezes estamos sem contactar a família várias semanas, sem nada de parecido sequer com um chuveiro, frios nocturnos e canícula insuportável diurna, comidas nem sempre com os standards gastronómicos… – existem experiências inolvidáveis.
Apenas alguns exemplos.
No segundo ano que estive na província de Neuquén, na Patagónia argentina, cheguei ao acampamento, a cerca de 150 km da povoação mais próxima, durante a noite (o diário da expedição foi já publicado n’O Primeiro de Janeiro).
Cerca das três da manhã e por motivos fisiológicos tive que deixar a tenda. Mal saí fui “assaltado” pela enormidade do céu estrelado que ao mesmo tempo me atraía e assustava. O céu parecia abarcar tudo, provocando quase uma sensação física de tão intenso e grande. Senti-me de uma pequenez extrema… Só pela vista deste céu a terrível viagem já havia valido a pena.
Durante o tempo que permaneci na Patagónia fui diversas vezes “atacado” pela beleza da paisagem, ao mesmo tempo inóspita e terrivelmente atraente; o nunca acabar da planície, o percebermos que somos tão pequenos…
Aliado a este lado atraente, que a maioria das pessoas facilmente entende e deseja, existe um outro – a camaradagem. Como referi, as condições de trabalho e de vida em expedições paleontológicas são as mais básicas que se pode imaginar. Apesar disso, surgem relações humanas de camaradagem e amizade que, noutros enquadramentos mais sofisticados, dificilmente poderiam nascer.
Em especial à noite, à volta de uma fogueira readquirem-se hábitos ancestrais esquecidos -contam-se histórias em grupo, fazem-se silêncios enquanto crepita o fogo, esquecem-se hierarquias académicas, ouvem-se pequenos desabafos pessoais.
Quis apenas relembrar e celebrar um dos aspectos envolvidos no processo científico da Paleontologia – o trabalho de campo.
Não vem nos relatórios nem nas publicações científicas. Não existem tratados nem compêndios que o analisem e sistematizem. Apesar de tudo isso tenho constatado que é das coisas que colocam um sorriso sincero na cara de cada paleontólogo – “Vais para o campo?”
Esquecem-se labutas diárias de obtenção de fundos, de preenchimento de formalidades burocráticas, da falta de perspectivas profissionais de futuro.
E vai-se…
P.S.:Este texto é um agradecimento pelo outro lado do trabalho científico a que me dedico.
E que, tendo a sorte de o ter, o gostaria de partilhar.
E existem tantas histórias, clássicas ou pessoais que envolvem o trabalho de campo…
(Publicado no jornal O Primeiro de Janeiro a 5/3/2007)
Todos os Nomes*
 Nomear e classificar são actos intrinsecamente humanos. Chamamos as coisas pelos nomes para as discriminar, mas esse acto acrescenta algo mais do que a mera nomeação da coisa.
Nomear e classificar são actos intrinsecamente humanos. Chamamos as coisas pelos nomes para as discriminar, mas esse acto acrescenta algo mais do que a mera nomeação da coisa.
“Não me chames nomes!”, dizia-me um colega de escola para não ser ofendido; “Isso não se diz!”, corrigia a minha mãe quando eu apelidava, com várias intenções, alguém de quem não gostava.
A classificação dos seres vivos, quer actuais quer os do registo fóssil, obedece a regras precisas com o objectivo de não gerar equívocos e mal-entendidos na comunicação científica.
Utilizando várias fontes online, todas baseadas na “bíblia” da nomenclatura dos animais – o ICZN (International Code of Zoological Nomenclature), dediquei algum tempo à pesquisa de nomes inusitados com que alguns animais têm sido nomeados.
Não poderia deixar de começar com uma personagem fóssil que tem estado ligada à minha vida profissional – o Apatosaurus louisae. Este dinossáurio saurópode americano foi dos primeiros a ser descoberto e montado numa exposição, nos finais do séc. XIX. O mecenas das escavações e trabalho científico, Andrew Carnegie, tinha como esposa a senhora Louise, sendo esta a musa inspiradora para o nome do grande animal.
Mas a homenagem dos paleontólogos não se ficou pela patroa; também o senhor Carnegie teve direito ao seu quinhão – Diplodocus carnegii – outro saurópode, sendo assim apelidado para preservar o nome do chefe. Não morder a mão que nos alimenta e abençoar o seu nome devia ser a moral da história…
Ainda no mundo dos dinossáurios de referir o Masiakasaurus knopfleri, um carnívoro descoberto em Madagáscar. A história do baptismo é conhecida mas foi-me contada na primeira pessoa por um dos autores, Catherine Foster. Justificou-me ela que ouviam a música dos Dire Straits, cujo vocalista é Mark Knopfler, quando descobriram o sáurio. Espero estar ouvir Amália da próxima vez que estiver a escavar, pensei eu na altura. E não é que existem pelo menos 13 animais com o nome “amalia”, entre aves, moluscos e insectos?
Outro dos ícones portugueses, o Pantera Negra, entra no nome científico de dois animais – Libellula eusebioi (uma libélula fóssil) e um ácaro das Filipinas, Dolicheremaeus eusebioi.
Aproxima-se o importuno Dia dos Namorados mas nada como relembrarmos o Bela eva, um gastrópode de Moçambique. Que nome bonito e adequado para a nossa cara-metade. Bem, a menos que ela seja uma malacologista – estudiosa de gastrópodes e bivalves – pois nesse caso não achará grande originalidade ao nosso piropo conjugal. Mas podemos sempre atirar-lhe com Amoraster – ouriço-do-mar do Miocénico da Austrália. Quem feio escuta, bonito lhe parece, pelo menos nesta situação.
Há 470 milhões de anos, quando fossilizou, o equinoderme primitivo Delgadocrinus oportovinum mal imaginava que teria o seu baptismo científico associado ao produto agrícola mais famoso de Portugal – o Vinho do Porto. Pois foi isto que a equipa liderada por um paleontólogo da UTAD fez; a justificação para a designação da nova espécie assenta na sua morfologia já que esta faz lembrar um cálice do néctar duriense, sendo a primeira parte do nome uma “vénia” ao geólogo Nery Delgado, responsável pelas primeiras recolhas de material.
Se os paleontólogos puderem ter algum apoio do sector vinícola, é ouro sobre…um Porto de Honra!
” ****, seu **** “, são nomes que não posso repetir e que apelidam o senhor do apito que corre de calções, com outros senhores que pontapeiam uma bola; mas quando “joga” o Taedia benfica – insecto pertencente à família Miridae e descrita por cientistas brasileiros, não me parece que haja muitos protestos, salvo se estivermos para os lados das Antas ou Alvalade.
O prémio para nome contraditório poderia perfeitamente ser atribuído à Boa canina – um réptil da ordem Serpentes; esta besta, que mata por estrangulamento, de boa não tem nada, pelo menos para as suas vítimas; e de canino, talvez apenas a fidelidade ao seus instintos.
Já o Turanogryllus mau – um grilo africano – pode invocar que tem a reputação manchada pelo nome científico que lhe calhou na rifa…

O mundo da Internet já serviu igualmente de inspiração ao “negócio” de alcunhar animais. A Proceratium google é uma formiga de Madagáscar, descrita em 2005 e, justificam os investigadores, tem a particularidade de saber procurar muito bem as suas presas, tal como o seu homónimo informático.
A lista de nomes é imensa, mas finalizo com Libellago finalis – uma libelinha.
 Assim, acho que ainda tenho tempo para agarrar La cerveza – uma traça norte-americana.
Assim, acho que ainda tenho tempo para agarrar La cerveza – uma traça norte-americana.
*Uma sincera homenagem à obra homónima de José Saramago.
(Publicado no jornal O Primeiro de Janeiro a 15/2/2007)


{kind=link}