


Uma Passagem para a Água
Este post faz parte de uma série cujo primeiro é O Poder dos Prótons.
Na fisiologia clássica, por volta da década de 20, quando se descobriu que a membrana plasmática das células era uma dupla camada lipídica, formulou-se a hipótese de que a água pudesse penetrar no meio intracelular através da própria membrana, seguindo forças osmóticas. Entretanto, uma série de pesquisadores, por meio de medidas biofísicas (que analisam potenciais de membrana), notaram que a permeabilidade de algumas membranas à água era 10 a 20 vezes maior do que a esperada caso a passagem ocorresse apenas através delas[1]. Previram, assim, a existência de um canal para passagem da água.
No início da década de 80, Peter Agre era um hematologista envolvido no estudo do fator Rh (o “positivo” ou “negativo” dos tipos sanguíneos) trabalhando no Instituto Nacional de Saúde dos EUA. Estava interessado em induzir a formação de anticorpos em coelhos sensibilizando-os com um peptídeo parcialmente purificado do fator Rh. Os coelhos reagiam fortemente produzindo anticorpos que, no entanto, não reagiam com o cerne da molécula de Rh mas sim, com uma outra proteína que acreditava-se ser um fragmento do grande polipeptídeo. Parecia uma contaminação do experimento. Mas essa proteína de 28 kilodaltons (kDa), tinha algumas propriedades estranhas. Não se corava com os procedimentos habituais e quando foram procurá-la na membrana de hemáceas, verificou-se que era extremamente abundante. Com aproximadamente 200.000 cópias por hemácea, era uma das proteínas mais comuns na célula! Nas palavras de Agre (descendente de imigrantes nórdicos) “era como se alguém, andando pelo norte desértico da Suécia, de repente, encontrasse uma cidade de 200.000 habitantes que não constasse em nenhum mapa”. Estudos subsequentes revelaram que a proteína tinha características de um canal pois dispunha-se ao longo da espessura da membrana. Uma série de elegantes experimentos revelou que ela era um canal de água sendo batizada com o sugestivo nome de aquaporina 1 (AQP1). Peter Agre recebeu o prêmio Nobel de Química em 2003 por sua descoberta (Figura abaixo). Orgulho do pai, que havia trabalhado com Linus Pauling, acabou abandonando a hematologia. (Veja a interessantíssima “palestra do prêmio” (em inglês, 45 min) no portal do Nobel).

Figura 1. Peter Agre. Prêmio Nobel de Química de 2003
As aquaporinas constituem uma família de, até o momento, algumas centenas de proteínas de membrana. Nos mamíferos já foram identificadas doze, sete das quais estão presentes no rim. Nos últimos anos, pesquisas têm explorado a seletividade e as funções multitransportadoras das aquaporinas. Isso levou a uma divisão do grupo em aquaporinas clássicas e aquagliceroporinas. Estas últimas também transportam passivamente glicerol e outros polióis bem como alguns solutos e, ao que parece, são filogeneticamente até mais antigas que as aquaporinas clássicas[2]. A AQP1 é uma proteína de membrana altamente permeável à água. Sua condutância é de aproximadamente 3 x 10^9 molec / subunidade / seg permitindo que uma molécula de água trafegue a ~0,02 cm/s [3]. A AQP1 forma 4 complexos na membrana celular (subunidades), cada um formando um canal de água independente (figura 2). Um quinto poro é formado no centro do complexo e há indícios de que possa conduzir íons, mas o transporte passivo de água através da membrana celular é mesmo a maior função fisiológica da AQP1 [4].

Figura 2. Foto de simulação do transporte de água por uma molécula de aquaporina. As subunidades são constituídas de túbulos que estão em cores diferentes. As moléculas de água (pequenos bumerangues azuis) permeiam a subunidade à direita (em dourado). Esta imagem ganhou o prêmio de melhor foto científica da revista Science™ na edição de 24 de Setembro de 2004 e pode ser encontrada aqui, juntamente com uma animação do transporte das moléculas de água.
É surpreendente o fato de que, mesmo transportando água tão eficientemente, as aquaporinas sejam impermeáveis aos íons. A razão disso seria a proteção do meio interior celular para que a maquinaria metabólica possa funcionar adequadamente e em “silêncio biológico” – o que pode até ser uma definição bonita de Saúde, mas teleológica demais para servir a nossos propósitos. Até o mais enxerido de todos íons, o próton e seu alterego H3O+, não consegue passar pelas reentrâncias do canal de água. Como isso se dá? Como isso ocorre se os prótons teleportam-se de um lugar para outro livremente na Mattrix aquática, materializando-se e evaporando em qualquer lugar bastando para isso um “fio de água” por onde possam passar…
É o que tentarei descrever no próximo e último post da série.
Referências Bibliográficas
[1] Finkelstein A: Water Movement Through Lipid Bilayers, Pores, and Plasma Membranes: Theory and Reality (Distinguished Lecture Series of the Society of General Physiologists). New York, John Wiley & Sons Inc, 1987.
[2] Nielsen S, Frokiaer J, Marples D, et al.: Aquaporins in the kidney: from molecules to medicine. Physiol Rev 82:205-244, 2002.
[3] Preston GM, Carroll TP, Guggino WB, et al.: Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 256:385-387, 1992.
[4] Zhu F, Tajkhorshid E, Schulten K: Theory and simulation of water permeation in aquaporin-1. Biophys J 86:50-57, 2004.
O Poder dos Prótons
Há um poder, silencioso e gigantesco que, entre outras coisas, é o responsável pela água na forma como a conhecemos no nosso planeta. Se a água é, por sua vez, a matriz de todas as formas vivas daqui, então tal poder também estará presente em cada ser vivo. A explosão de vida da Terra teve que lidar com ele para que pudesse florescer e, de fato, quando nos escapa esse controle, uma fera enjaulada foge e a vida diminui ou mesmo acaba. Podemos chamar isso de “doença”. Nos próximos posts vou tentar contar a história do Poder dos Prótons e seus desdobramentos na fisiologia e fisiopatologia dos seres vivos, em especial, dos humanos.
O Hidrogênio poderia, para todos os efeitos, ser considerado um elemento-traço. Para se ter uma ideia, sua concentração é 3,5 milhões de vezes menor que a do Sódio nos fluidos orgânicos. No soro, ela é da mesma ordem de grandeza do Molibdênio (20 nM/L) e bem menor que concentrações de elementos como Zinco (15 mM/L), Cobre (20 mM/L) e Selênio (1 mM/L) [1]. Como pode o Hidrogênio com uma concentração de 0,000 000 040 M/L ou 40 nM/L influenciar tão decisivamente os processos biológicos? Quais as razões que levam o íon hidrogênio a desempenhar um papel tão importante?
A primeira delas é, sem dúvida, a água. Todos os fenômenos biológicos ocorreram e ocorrerão sempre em soluções aquosas desde que a estrutura celular que caracteriza os seres vivos neste planeta seja mantida. A água tem, como se sabe, estranhas propriedades quando comparada a outros hidretos da família do Oxigênio (a família 6A na tabela periódica – Enxofre, Selênio, Telúrio e Polônio): altos pontos de fusão e ebulição, alta constante dielétrica (conduz correntes bem), alta tensão superficial (forma gotas muito resistentes), entre outras (Tabela 1).
|
Grupo |
3A |
4A |
5A |
6A |
7A |
|
B2H6 : -92.5 |
CH4 : -164 |
NH3 : -33.4 |
H2O : +100 |
HF : -87.7 |
|
| H2S : -60.7 | |||||
| H2Se : -41.5 | |||||
| H2Te : -2 |
Tabela 1. A água tem um ponto de ebulição anormalmente elevado quando comparado por extrapolação em relação aos hidretos do grupo 6A (Oxigênio, Enxofre, Selênio e Telúrio). Os hidretos dos outros grupos são mostrados para comparação (3A Boro, 4A Carbono, 5A Nitrogênio e 7A Flúor). (Retirado daqui)
A grande maioria das estranhas propriedades da água provém da habilidade que ela tem de formar pontes de hidrogênio. Como se sabe, a ligação do Oxigênio aos dois átomos de Hidrogênio dá à molécula de água o formato de um “V” com um ângulo de 104,5o. Essa ligação é formada por um par de elétrons compartilhado (a chamada ligação covalente). O Oxigênio, no entanto, atrai o par de elétrons para bem perto de si (maior eletronegatividade) e transforma a molécula de água em um minúsculo imã com seus 2 pólos: o positivo (Hs) e o negativo (O). Quando um átomo de H, carregado positivamente, fica preso entre dois átomos de O, que têm cargas negativas, ele passa a funcionar como uma ponte entre os dois. Essa atração é 90% eletrostática e 10% covalente (há evidências de que eles podem realmente dividir orbitais, fazendo com que não consigamos saber qual ligação é a covalente), e cada molécula de água tem o potencial de fazer 4 pontes. Além disso, pode-se dizer que a ponte de hidrogênio é cooperativa isto é, uma vez formada a primeira ponte, a molécula torna-se mais apta a formar outras pontes e o fenômeno se dissemina. O contrário também é verdadeiro, ou seja, é mais difícil romper a primeira ponte, sendo que a energia necessária para romper as subsequentes vai, progressivamente, ficando menor. Isso transforma a água em um “sincício”, tipo um tecido tridimensional; daí a poderosa força de atração intermolecular que dota a água de seus elevados pontos de ebulição, fusão e tensão superficial. A formação das pontes de hidrogênio faz com que a entalpia (energia) do sistema fique mais negativa e a entropia (grau de desorganização) menos positiva ou, trocando em miúdos: a água se organiza!. Organiza-se com baixo custo, o que significa que é difícil tirá-la desse estado de organização e, na dependência da temperatura e pressão, temos a explicação de muitas das suas “estranhas” propriedades. Ou seja, se a água fosse apenas H2O seria um gás na faixa de temperatura que permite que seres vivos sobrevivam. No estado líquido, ela está na forma de (H2O)n, com n→∞, constituindo mais pontes de H que qualquer outro solvente e com quase tantas pontes quanto ligações covalentes. Esse sistema é interessante pois pode rearranjar-se rapidamente frente a estímulos como solutos ou alterações de temperatura, como uma rede tridimensional capaz de mudar o tamanho de seus “buracos” de acordo com o tamanho dos “peixes” que nela caem.
Além disso, a água demonstra uma extremamente baixa mas mensurável capacidade de formar íons. Ioniza-se devido a flutuações do campo magnético de seu dipolo causadas por vibrações complexas de sua estrutura molecular e também por posicionamentos espaciais favoráveis de suas pontes de H. Após a ionização, o H+ não permanece livre muito tempo (menos que 1% do tempo total) e é hidratado para formar o hidroxônio, oxônio ou hidrônio (H3O+). Os três átomos de H da molécula são equivalentes na ligação com o O e organizam as pontes com as moléculas de água adjacentes. O H3O+ exibe uma mobilidade frente aos campos elétricos de testes que é absurdamente maior que a prevista para um cátion monovalente (como o Lítio, por exemplo, que apresenta inclusive uma forma de hidratação semelhante). Qual seria a explicação para esse fenômeno?
Um próton pode viajar em uma solução aquosa de duas maneiras: uma, chamada hidrodinâmica, em que o H3O+ difunde-se como uma molécula comum abrindo caminho pelo meio aquoso; outra, chamada prototrópica e essa merece uma explicação. Em 1905 [2], foi sugerido que a transferência do próton poderia ocorrer por uma “rede” de pontes de hidrogênio, um processo que envolveria uma série periódica de polimerizações da água entre H9O4+ (cátion de Eigen) e o H5O2+(cátion de Zundel), conhecido como efeito Grotthuss [3], proton-wire ou proton-jumping, ou ainda water-wire. Isso só é possível, devido à efêmera duração da ponte (cada ponte tem uma duração média de 10 psec, sendo que 1 picosegundo = 10-12s) e à facilidade com que a água as forma e “re-forma”, como vimos. Dizemos então, que a alta condutividade do próton em meio aquoso ocorre devido a “mudança de identidade das moléculas de água que participam das pontes” pois os núcleos de H vão passando de uma para outra conforme esboça a figura 2, sendo que o que entra, é diferente daquele que sai.
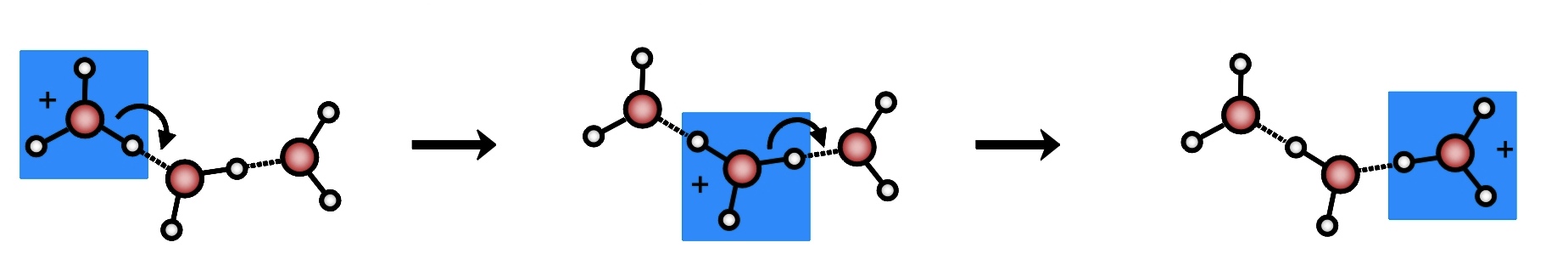
Figura 2. Esquema simplificado do efeito Grotthuss
Isso funciona parecido com o hipnotizante Pêndulo de Newton, na figura abaixo.

Pêndulo de Newton (fonte Wikipédia)
Mas…
O íon hidrogênio tem uma densidade de carga altíssima e isso ocorre porque ele é um cátion-anão (monovalente no qual não existem elétrons circulantes) sendo seu raio 105 vezes menor que qualquer outro. Por essa razão, tem um grande campo magnético ao seu redor. Assim, mesmo estando presente em concentrações baixíssimas nas soluções e devido sua alta mobilidade, o H3O+ é um “pentelho iônico” e pode afetar a conformação de proteínas, ácidos nucléicos e membranas biológicas, bastando para tal, que haja uma “brecha” eletrostática (cargas) na solução. Esse é o “poder dos prótons”, capazes de alterar todos os processos biológicos, portanto. Fenômenos semelhantes ocorrem com a hidroxila OH– entretanto, sua mobilidade nas soluções tem sido bem menos estudada. Pesquisas recentes [4] demonstram que sua mobilidade também é anormalmente alta para um ânion de seu porte e, portanto, mecanismos semelhantes ao do H3O+ podem também ocorrer. Tanto o H+ como o OH– são gerados pela destruição e destruídos pela formação de uma molécula de água. Se considerarmos que a água é, em ordem de grandeza, a mais concentrada substância dos sistemas vivos (55,3M – o que é 400 vezes mais concentrado que o sódio), e que ela provê uma fonte simplesmente inesgotável para esses íons, entenderemos porque eles se comportam de maneira tão diferente de outros íons de carga e massa semelhantes.
Com toda essa mobilidade e onipresença, é natural esperar que prótons penetrem as células com facilidade, podendo fazê-lo através da membrana ou por meio de canais, específicos ou não, para seu transporte. Tal fluxo é tão fundamental quanto o de água para a célula, pois está ligado à produção da energia que sustenta o metabolismo celular. Mas como evitar então, que o transporte de água em abundância através da membrana não cause uma catástrofe iônica na célula? Como evitar que os “pentelhos iônicos” dos H3O+ saturem as cargas das proteínas e as deformem fazendo com que percam sua função e, assim, parem o funcionamento da maquinaria celular?
É o que mostraremos no próximo post.
Referências Bibliográficas
[1] Tietz N: Clinical Guide to Laboratory Tests, 3rd ed. Philadelphia, W. B. Saunders, 1995
[2] Decoursey TE: Voltage-gated proton channels and other proton transfer pathways. Physiol Rev 83:475-579, 2003.
[3] Agmon N: The Grotthuss mechanism. Chem Phys Lett 244:456-462, 1995
[4] Tuckerman ME, Marx D, Parrinello M: The nature and transport mechanism of hydrated hydroxide ions in aqueous solution. Nature 417:925-929, 2002

