


A Febre Hemorrágica do Ebola
Em 1976, no antigo Zaire, hoje República Democrática do Congo (RDC), uma estranha e desconhecida doença caracterizada por febre alta, hemorragias na pele e nos órgãos internos, começou a acometer os moradores de uma pequena cidade ribeirinha chamada Yambuku. A mortalidade chegou a quase 90% e chamou a atenção das autoridades. Após auxílio internacional, um grupo de pesquisadores descobriu que o agente causador era um vírus filamentoso da mesma família do Marburg descrito na Alemanha, quase uma década antes. O vírus foi batizado como Ebolavirus (pronuncia-se ébola) dado que a doença já era conhecida como Febre Hemorrágica do Ebola, segundo o rio que banhava a cidade, tributário do Mongala que, por sua vez, desemboca no grande rio do Congo.
No mesmo ano, nas cidades de Maridi e Nzara, Sudão do Sul, a mais de 1000 km em linha reta ao norte de Yambuku, uma epidemia com características muito semelhantes também eclodiu. Pesquisadores encontraram uma variante do mesmo vírus do Ebola nos pacientes acometidos. Outras epidemias surgiram no final da década de 70, não só na África, o que, ao fim e ao cabo, permitiu a identificação de 5 sorotipos da família Ebola: 1. Bundibugyo ebolavirus (BDBV); 2. Zaire ebolavirus (EBOV); 3. Reston ebolavirus (RESTV); 4. Sudan ebolavirus (SUDV); 5. Taï Forest ebolavirus (TAFV). O RESTV foi encontrado em macacos importados de Mindanao, nas Filipinas, para Reston, Virginia, nos Estados Unidos e não é considerado causador de doença humana.
O comportamento epidemiológico estranho do Ebola não parou por aí. Após ter aparecido na África em 1976 – 1979, ele simplesmente sumiu das estatísticas até 1994. Acredita-se que permaneceu oculto, circulando em seus reservatórios naturais. Um reservatório natural de um patógeno é um hospedeiro, animal ou inseto, que por não apresentar as características da doença, “convive” com o agente de forma pacífica. No Brasil, tatus são reservatórios para doença de Chagas e morcegos podem abrigar o vírus da raiva. O problema é que, no caso do Ebolavirus, esse reservatório, até hoje pelo menos, não foi confirmado, suspeitando-se de roedores e morcegos já que seu primo, o Marburg, tem como reservatório um morcego de caverna (o Rousettus aegyptiacus).
A Febre Hemorrágica do Ebola é o protótipo de uma zoonose. Uma zoonose é uma doença transmitida por animais (e.g. Raiva), e também não foram encontrados vetores para a doença. Um vetor é um bicho (pode ser animal ou inseto) que transmite determinada doença (e.g. o mosquito da Dengue, Aedes aegypti). O comportamento esquivo das epidemias poderia ser explicado por situações que alterariam o ecossistema dos reservatórios proporcionando altos índices de infecção nos humanos de tempos em tempos.
Quadro Clínico
O período de incubação da doença é de 4 a 10 dias (variando de 2 a 21). É, logo de início, uma doença sistêmica (como são por exemplo, os casos de infecção generalizada por bactérias – sepse -, as vasculites de causas imunológicas, a pancreatite aguda necro-hemorrágica, grandes queimados e politraumatismos extensos), o que dificulta seu tratamento e aumenta sua gravidade. O paciente sente-se extremamente indisposto, com dor abdominal, náuseas e vômitos (que podem conter sangue), diarreia (com ou sem sangue), falta de ar, tosse, pressão baixa, dor de cabeça, confusão mental e coma. Podem ocorrer hemorragias cutâneas e viscerais e a pele costuma descamar para cicatrizar ad integrum nos sobreviventes. Um quadro chamado de coagulopatia intravascular pode ocorrer e é, em geral, fatal. Virtualmente, todos os vasos sanguíneos do organismo podem sangrar e o quadro é realmente dramático. Tal problema não é, infelizmente, incomum em casos graves causados por outras doenças e não é uma exclusividade do Ebola. A Dengue, por exemplo, é a febre hemorrágica mais comum do planeta.
Mortalidade
A virulência do Ebola parece depender do sorotipo causador. A mais letal parece ser a EBOV seguida pela SUDV, com 60–90% e 40-60% de case-fatality (porcentagem de pessoas que contraíram a doença e que vieram a falecer dela, direta ou indiretamente), respectivamente, o que é muito. Devemos lembrar, entretanto, que tais epidemias ocorreram em locais onde os cuidados médicos estavam longe de ser ideais o que pode falsear as estatísticas. De qualquer forma, é uma doença muito grave.
Modo de Infecção
O Ebolavirus parece penetrar no organismo humano através de superfícies mucosas, lesões cutâneas, ferimentos ou pelo sangue. Muitas das infecções causadas em humanos, em especial o grande contingente de agentes da saúde, parecem ter ocorrido pelo contato direto com pacientes infectados. Partículas de RNA vital foram encontradas no esperma e secreções vaginais de pacientes, bem como em outros fluidos como secreções nasais. Ferimentos acidentais por agulhas e material contaminado são importantes rotas de infecção. As epidemias africanas da década de 70 foram atribuídas ao consumo de morcegos e macacos. Não é confirmada, até o momento, a transmissão por aerossóis.
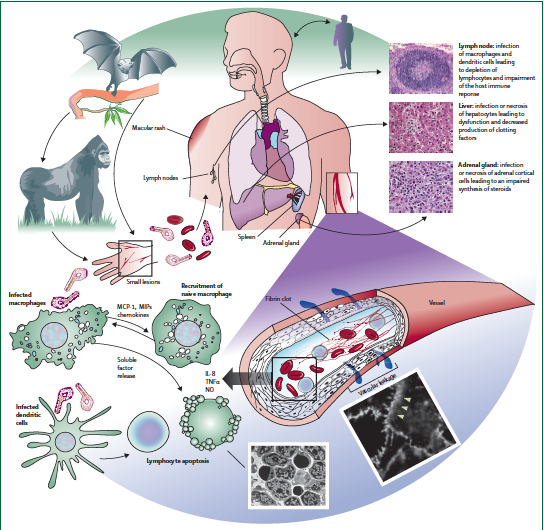
Copyright The Lancet, 2011
Precauções e Tratamento
O isolamento dos pacientes é uma eficiente medida de proteção já que o tratamento específico ainda é objeto de pesquisas. Medidas de suporte precoces como hidratação e cuidados intensivos são muito importantes e fazem grande diferença na mortalidade. Entretanto, há que se chamar a atenção para o caso dos dois agentes sanitários americanos recentemente submetidos a uma terapia experimental para o tratamento da doença contraída em campo. Kent Brantly e Nancy Writebol contraíram a Febre Hemorrágica do Ebola na Libéria trabalhando em uma organização cristã de auxílio às vítimas da doença. Apesar do protesto absurdo de alguns sobre a repatriação de americanos infectados, o fato é que, baseados em um relato de caso no qual o soro de pacientes sobreviventes conseguira reverter a doença, um consórcio de pesquisadores conseguiu desenvolver uma solução com anticorpos monoclonais contra determinados antígenos imunogênicos do vírus e curar os dois missionários, fato amplamente divulgado na mídia.
Futuro
Não é impossível que a Febre Hemorrágica do Ebola desembarque no Brasil. Contam a nosso favor, seu modo peculiar de transmissão, seus possíveis reservatórios (ausentes aqui?), nossa experiência em tratar a dengue hemorrágica e nosso sistema de saúde, que se não é a segunda maravilha do mundo, é bastante superior e estruturado comparado ao da maioria dos países africanos. Contra nós, pesa nossa eterna ineficiência sanitária e as grandes conglomerações nas cidades.
De vez em quando, uma epidemia qualquer com “potencial” de extermínio da humanidade “aporta” neste blog. Gosto de citar como exemplo minha pior experiência com tais epidemias que foi com a Síndrome de Weil, a forma hemorrágica da leptospirose humana, na epidemia ocorrida em São Paulo no final dos anos 90. Naquela época, de enchentes, diga-se de passagem, a doença transmitida pela convivência promíscua com roedores, atingiu em cheio a parte mais pobre da população. Ver homens, mulheres e crianças em macas sangrando por quase todos os orifícios foi das minhas piores vivências que tive como médico. Passou. Estamos aqui. No caso do Ebola, já temos até o caminho a seguir para conseguir a cura. Resta saber se as instituições envolvidas na descoberta do “soro mágico” contra a doença possibilitem sua oferta para quem mais necessita. Isso sim, a possibilidade de me decepcionar uma vez mais com a espécie humana, me dá um medo desgraçado.
Bibliografia
![]() Feldmann, H., & Geisbert, T. (2011). Ebola haemorrhagic fever The Lancet, 377 (9768), 849-862 DOI: 10.1016/S0140-6736(10)60667-8
Feldmann, H., & Geisbert, T. (2011). Ebola haemorrhagic fever The Lancet, 377 (9768), 849-862 DOI: 10.1016/S0140-6736(10)60667-8
![]() Peters, C., & LeDuc, J. (1999). An Introduction to Ebola: The Virus and the Disease The Journal of Infectious Diseases, 179 (s1) DOI: 10.1086/514322
Peters, C., & LeDuc, J. (1999). An Introduction to Ebola: The Virus and the Disease The Journal of Infectious Diseases, 179 (s1) DOI: 10.1086/514322
![]() Mupapa, K., Massamba, M., Kibadi, K., Kuvula, K., Bwaka, A., Kipasa, M., Colebunders, R., Muyembe‐Tamfum, J., & , . (1999). Treatment of Ebola Hemorrhagic Fever with Blood Transfusions from Convalescent Patients The Journal of Infectious Diseases, 179 (s1) DOI: 10.1086/514298
Mupapa, K., Massamba, M., Kibadi, K., Kuvula, K., Bwaka, A., Kipasa, M., Colebunders, R., Muyembe‐Tamfum, J., & , . (1999). Treatment of Ebola Hemorrhagic Fever with Blood Transfusions from Convalescent Patients The Journal of Infectious Diseases, 179 (s1) DOI: 10.1086/514298
DEK – Olhar e Ver Espelhos

Xadrez no Espelho
O que é “olhar”? Literalmente, olhar é “dar uma olhada”, um golpe d’olhos. É o movimento conjugado dos globos oculares em direção a algo ou alguém. Se, por um lado, quem não olha fatalmente não vê, como é sabido de todos, o inverso é mesmo possível, qual seja olhar de fato, sem nada ver. Por quê? “Ver”, assim como seu correlato “enxergar”, parecem conferir algo de interpretativo ao ato mesmo de olhar. A etimologia de “enxergar” é classificada como “incerta” pelo Houaiss e pelo Dicionário Etimológico da Língua Portuguesa, o que nos permite e(spec)ular (as razões desses parênteses serão esclarecidas abaixo). Em castelhano temos a palavra envidia que vem do latim invidere, sendo composta por “in-“ “pôr sobre”, “ir para” e “videre”, o próprio “ver”. Envidia significa, portanto, algo como “deitar o olhar sobre” e seria um étimo possível para nosso “enxergar”. Por outro lado, em latim ainda temos a palavra insecare, primeira pessoa do verbo inseco, que significa “cortar, divisar”. Foi sugerido [1] que a relação entre “cortar” e “saber”, que é também dada no vocábulo derivado do grego “análise”, pudesse ter originado o “enxergar”. Incerto, de qualquer forma, mas plausível e certamente aprovado ao menos por Michel Foucault[2].
E o que é “ver”? A origem é o latim videre como vimos e que, por sua vez, vem de uma raiz indoeuropeia *weid-, comum, veja só, à palavra grega ειδος (eidos = aparência, imagem) tão cara à Platão e que originou as palavras “androide”, “antropoide”, “ginecoide” e tantas outras com o significado de “assemelhado a” ou “na forma de”. Interessante também o fato de que, em bom inglês, tal raiz tenha originado wisdom (sabedoria), wise (sábio), wizard (mago), todas palavras que de certa forma designam a capacidade que alguém tem de “ver mais longe”.
Peculiar é o termo species que também significa “aparência”, “a(spec)to” (calma, já chegamos lá) e “visão” e deriva de uma raiz (spec) (pronto!) que significa “olhar, ver”, raiz essa que pode ser encontrada também em palavras como speculum, que não significa apenas “espelho”, mas também é o nome que se dá a um instrumento médico utilizado para ver “interiores corpóreos”, muito utilizado em ginecologia (aliás, uma das “e(spec) ialidades” médicas); spectrum, “imagem”, “fantasma”; specimen, “exemplo”, “signo”; spectaculum, “espetáculo”. Raiz que, segundo Giorgio Agamben [3], se desdobra numa dialética bastante interessante. Species foi utilizado para traduzir para o latim o termo filosófico eidos (acima), derivando seu sentido para as ciências da natureza (espécie animal ou vegetal) e para o comércio, significando “mercadoria” e, mais tarde, o próprio dinheiro. Ainda segundo Agamben
“especioso” significa “belo” e, mais tarde, “não verdadeiro”, “aparente”. “Espécie” significa o que torna visível e, mais tarde, o princípio de uma classificação de equivalência. Causar espécie significa “assombrar, surpreender” (em sentido negativo); mas que indivíduos constituam uma espécie nos traz segurança” [4] (itálicos no original).
A espécie é, então, a imagem de uma coisa que se mostra ao olhar mas que, ao mesmo tempo, precisa ser fixada na própria coisa para se constituir em uma identidade. Por isso, a fórmula de Agamben é tão promissora: “especial” é o ser cuja essência coincide com seu dar-se a ver, com sua espécie. Quando alguém diz que somos especiais, tal afirmação pode constituir-se num elogio de autenticidade, mas também numa crítica de impessoabilidade ou mesmo de insubstancialidade. “Só personalizamos algo – referindo-o a uma identidade – se sacrificamos a sua especialidade” – diz Agamben.
E a coisa toda fica bem mais interessante quando observamos nossa própria “espécie” refletida num espelho. Isso porque o espelho é o locus da descoberta de que nossa “espécie”, nosso imago, não nos pertence. E “entre a percepção da imagem e o reconhecer-se nela há um intervalo que os poetas medievais denominavam amor“[5]. O espelho de Narciso é essa experiência. “Se eliminarmos esse intervalo ou o prolongarmos indefinidamente, a imagem é interiorizada como “fantasma”, e o amor recai na psicologia”[5], metáfora para patologização do Eu.
Nosso olhar seria então um meio pelo qual construimos um mundo e também reconhecemos os sujeitos que nele habitam. Entretanto, ao voltar-se sobre si e nos submeter ao escrutínio de seu recorte, um certo cuidado é preciso. Para que não comecemos a ver fantasmas onde eles jamais existiram.
[1] Takata, R. Personal communication.
[2] Foucault, M. Microfísica do poder. Rio de Janeiro: Graal, 1984. “É que o saber não é feito para compreender, ele é feito para cortar.” p. 28.
[3] Agamben, G. Profanações. Boitempo, 2007. Tradução e apresentação de Selvino J. Assmann. p. 52.
[4] Idem. p. 54.
[5] Idem. p. 53.
Transplante de Fezes
Parece* que nascemos mesmo livres, ao menos dos germes. Estéreis ou, como os cientistas gostam de dizer, gnotobióticos. Logo após o nascimento, entretanto, no período neonatal, somos já colonizados. O tipo de colonização é bastante influenciado pelo contato materno mas é, sob alguns aspectos, uniforme. Há mais de cinquenta filotipos de bactérias mas apenas quatro nos adotaram como lar. São eles os Firmicutes, os Bacteroidetes, as Actinobacterias e as Proteobacterias (parece Game of Thrones, né?). Mas, por que só quatro? Essa especificidade sugere uma interação de poderosas forças seletivas que permitiram eliminar algumas e manter outras espécies em co-evolução.
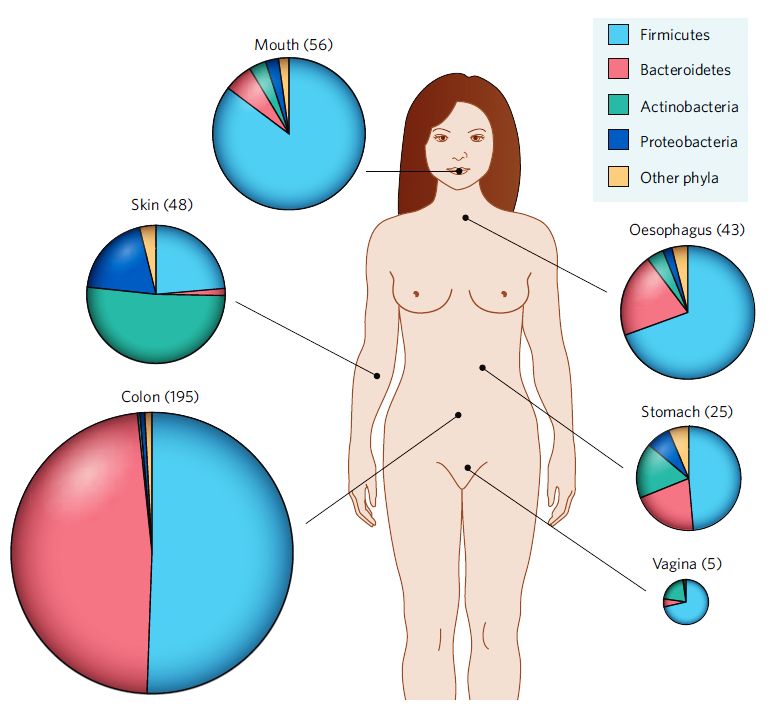
Fig. 1 – Esquema de distribuição dos filos bacterianos em um ser humano saudável. A área de cada setor do gráfico está relacionada ao número de diferentes espécies bactérias do filo no determinado sítio do hospedeiro. (copiado com permissão do Meio de Cultura)
Apesar de seguirmos esse padrão geral, cada ser humano parece ter uma combinação própria de bactérias desses filos, tornando a microbiota quase uma assinatura microbiológica de cada pessoa. As regras que governam a co-existência de humanos e bactérias de acordo com esse modelo ainda não são bem elucidadas, mas o fato é que alguns estudos em animais de laboratório gnotobióticos mostraram que tais bactérias não são fundamentais para o desenvolvimento normal desses bichos. Por outro lado, a gnotobiose não ocorre de forma espontânea na natureza. Então, parece realmente que obtemos alguma vantagem de nos deixar ocupar. No nosso caso específico, a microbiota que nos habita facilita a incorporação de vitaminas e nutrientes, ajuda no desenvolvimento e manutenção da integridade dos tecidos, em especial da mucosa intestinal, além de estimular em muitos aspectos as defesas contra invasores causadores de doenças, para citar alguns exemplos. Um desses invasores tem tirado o sono de médicos e microbiologistas: o Clostridium difficile. Vamos conhecê-lo mais de perto.
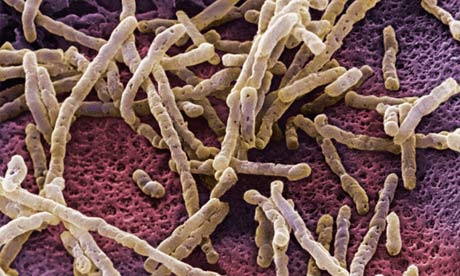
Fig. 2 – Fotografia de microscopia de população de Clostridium difficile no intestino sem microbiota nativa. (Copiado do The Guardian Foto de Dr David Phillips/Getty Images). Clique na foto para ver o original.
O Clostridium difficile é um bacilo (forma de bastão) anaeróbio obrigatório (não utiliza oxigênio), gram-positivo, que faz parte da microbiota intestinal e produz duas toxinas (A e B). A toxina A é uma enterotoxina que causa aumento da permeabilidade intestinal e secreção de fluidos, enquanto a toxina B é uma citotoxina que causa intensa inflamação dos cólons. Por isso, o clostrídio causa uma colite que chamamos de pseudomembranosa devido ao seu aspecto macroscópico à colonoscopia. O Clostridium difficile pode provocar até 20% dos casos de diarreia associada a antibióticos e é uma das causas mais comuns de diarreia adquirida em hospitais, em geral, em pacientes já debilitados o que além de dificultar o tratamento, aumenta o número de complicações. O tratamento para a colite por C. difficile consiste na administração oral de antibióticos (metronidazol ou vancomicina). Além disso, a taxa de recidivas é muito alta (15 a 26% dos pacientes). Nenhum tratamento efetivo contra as recorrências está disponível e o que temos é a prescrição de vancomicina prolongadamente. Isso, além de diminuir progressivamente sua eficácia, pode também ser a causa do problema. O que fazer?

Foi quando alguém teve a brilhante ideia que dá título a esse texto.
Ora, se o problema é um desbalanço na microbiota intestinal, vamos tentar restabelecê-la. Inicialmente, foram tentadas bactérias como os próbióticos (lactobacilos, etc). Não deu certo. Quando se tem pacientes morrendo e não se dispõe de terapias adequadas, a necessidade cria alternativas, muitas vezes desesperadas ou mesmo inusitadas.
“Por que não passamos as bactérias de um indivíduo saudável para um doente?” – pensou algum médico desesperado. “Sim. Mas como faremos isso?” – respondeu seu amigo pragmático. “Ora, infundimos um lavado de fezes do intestino do saudável para o doente… Simples!”. Foi o que fez o grupo de pesquisadores holandeses liderados por Els van Nood, citados abaixo. Por mais nojento que possa parecer, a coisa funcionou. E muito bem. O estudo teve que ser interrompido em sua análise interina pois, com apenas 43 pacientes randomizados, foi possível demonstrar uma nítida melhora no grupo tratamento com 16 pacientes. Qual tratamento? O intestino dos indivíduos com clostrídio era lavado e após isso, infundia-se, por meio de uma sonda nasogástrica, 500 ml de solução constituída de material fecal de um doador saudável (ver metodologia original no final do texto). Paralelamente a isso, foi tentado o tratamento convencional com vancomicina e ainda, uma mistura dos dois. A figura 3 mostra a taxa de cura de cada tratamento.
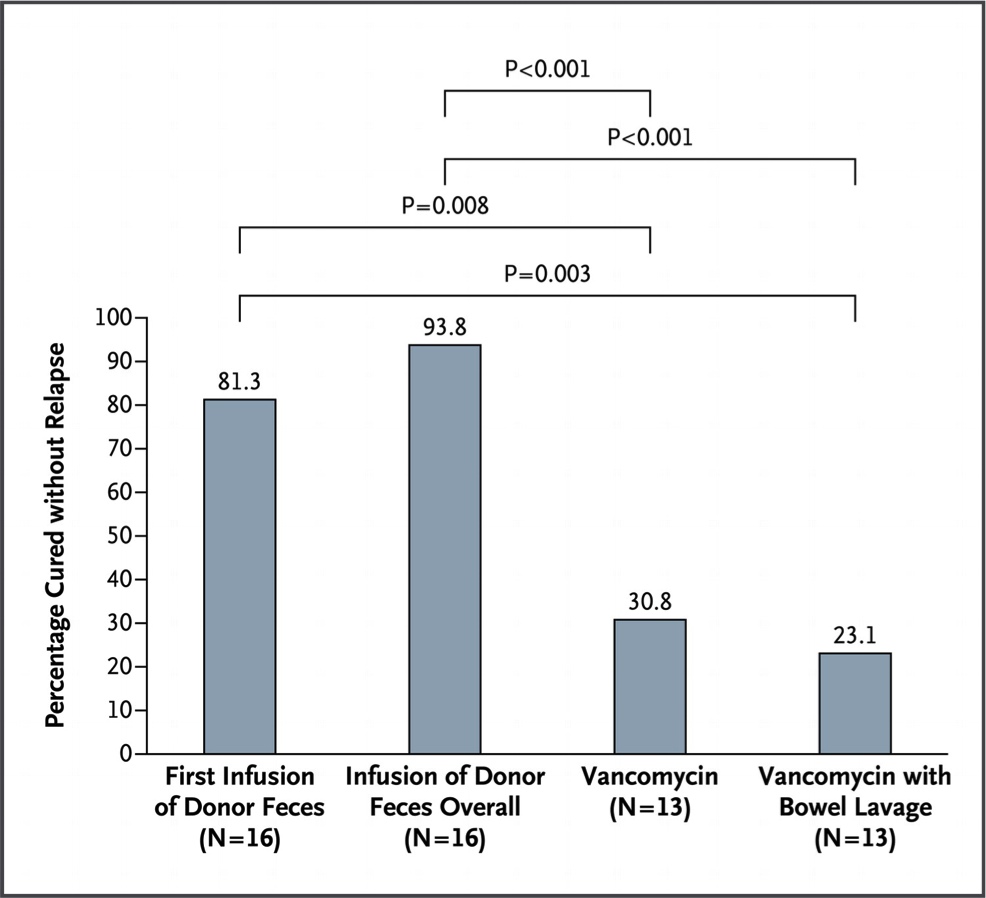
Fig. 3. Gráfico mostrando a taxa de cura de acordo com os tratamentos instituídos com as respectivas significâncias (p) de cada grupo comparado com os outros. Para mais explicações, ver o texto.
O primeiro grupo, representado na coluna mais a esquerda, mostra a taxa de cura com uma infusão única. Se isso não resolvesse, era tentada uma nova infusão com fezes de um doador diferente. Esses resultados são agrupados na segunda coluna do gráfico da figura 3. A vancomicina sozinha ou associada às infusões parece realmente ser inferior. Interessante também avaliar a diversidade da microbiota dos pacientes após as infusões.
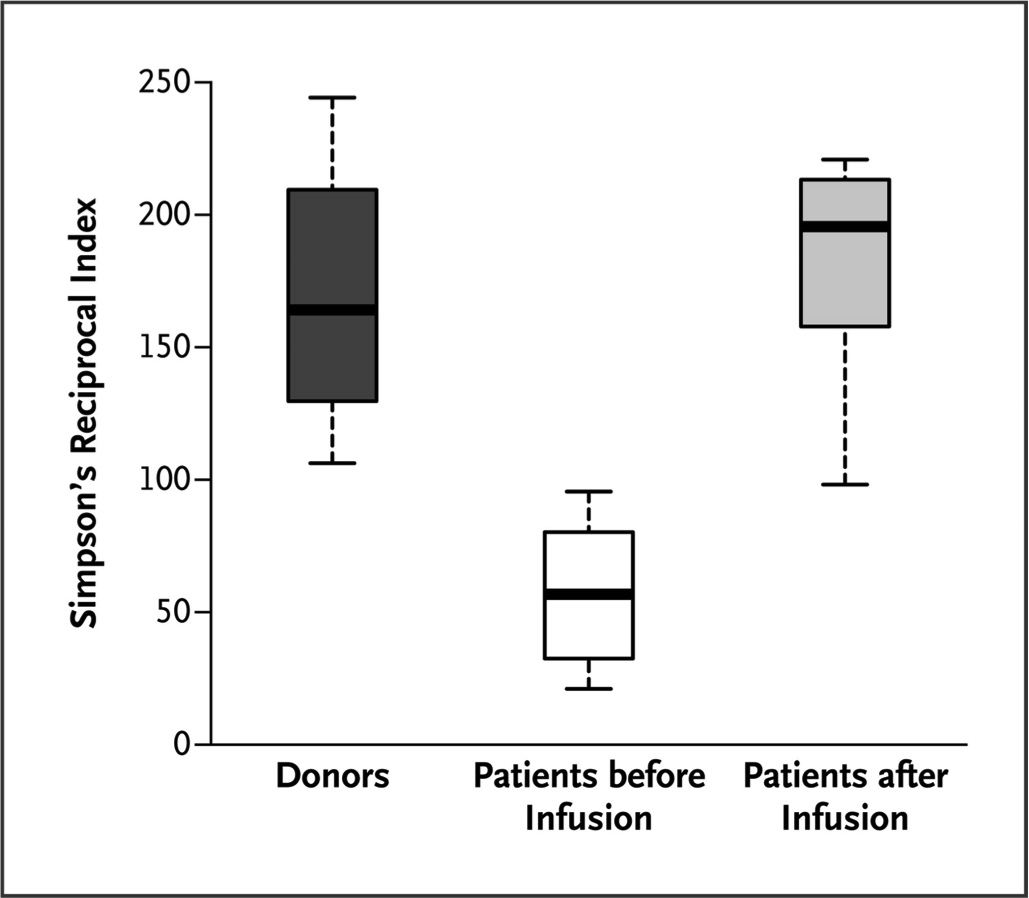
Fig. 4. Gráfico mostrando a variação da microbiota intestinal nos pacientes antes e após a infusão de fezes, comparando com os doadores de acordo com o índice de recíproco de Simpson. Para mais explicações, ver o texto.
A microbiota intestinal foi avaliada por análise de DNA usando microarray filogenético (Human Intestinal Tract Chip – HITChip) e a diversidade das comunidades bacterianas antes e depois da infusão de fezes usando uma escala chamada de Simpson’s Reciprocal Index of Diversity, que vai de 1 a 250, com os maiores valores indicando maior diversidade. É nítida a mudança após a infusão, ficando os pacientes com uma microbiota tão diversa quanto a dos doadores.
A conclusão é que o tal transplante de fezes funciona para o tratamento de infecções recorrentes pelo Clostridium difficile. Dito isto, podemos começar a imaginar algumas coisas. Muitas afecções atuais, que vão desde a obesidade até doenças cardíacas passando por fibromialgia, pancreatite e autismo, estão sendo atribuídas a alterações da microbiota (figura 5).
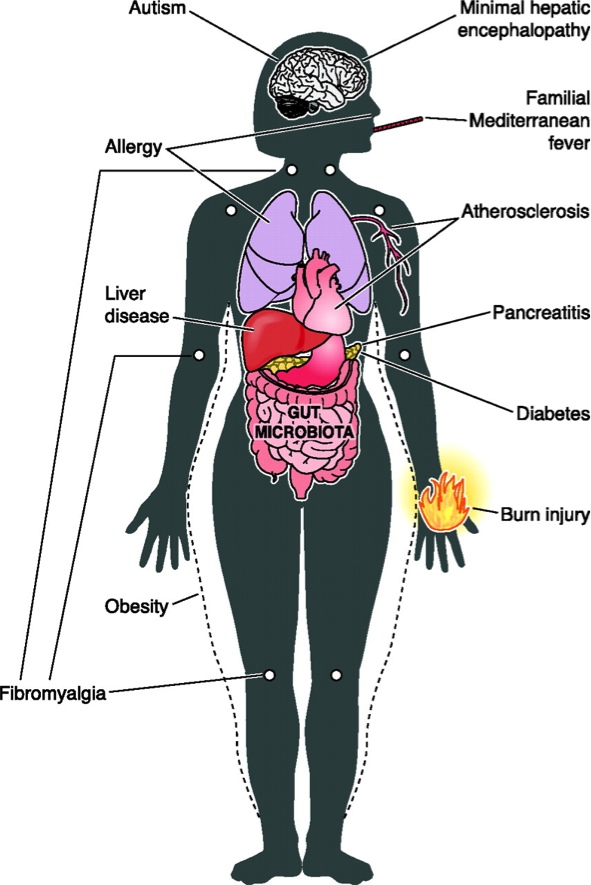
Fig. 5 – Esquema de possiveis doenças e/ou complicações associadas a alterações da microbiota intestinal. Clique na foto para ver o original.
Será que os futuros tratamentos vão ser…? Sei lá. Melhor nem pensar… Espero que alguém imagine uma cápsula deglutível que nos traga colonizadores saudáveis e nos livre de tais procedimentos pouco aprazíveis. Seria o caso de instituirmos um tipo de microhiperneocolonialismo do bem? Sim, porque a abordagem convencional de partir pra porrada e dar “veneno” para as bactérias, dividindo-as entre boas e más, parece não surtir mais efeito (como em tudo, aliás).
*Atualizado em 23/06/2013
Referências
Blaser, M., & Falkow, S. (2009). What are the consequences of the disappearing human microbiota? Nature Reviews Microbiology, 7 (12), 887-894 DOI: 10.1038/nrmicro2245
van Nood, E., Vrieze, A., Nieuwdorp, M., Fuentes, S., Zoetendal, E., de Vos, W., Visser, C., Kuijper, E., Bartelsman, J., Tijssen, J., Speelman, P., Dijkgraaf, M., & Keller, J. (2013). Duodenal Infusion of Donor Feces for Recurrent New England Journal of Medicine, 368 (5), 407-415 DOI:10.1056/NEJMoa1205037
Apêndice
Infusion of Donor Feces – methodology
Donors (>60 years of age) were volunteers who were initially screened using a questionnaire addressing risk factors for potentially transmissible diseases. Donor feces were screened for parasites (including Blastocystis hominis and Dientamoeba fragilis), C. difficile, and enteropathogenic bacteria. Blood was screened for antibodies to HIV; human T-cell lymphotropic virus types 1 and 2; hepatitis A, B, and C; cytomegalovirus; Epstein–Barr virus; Treponema pallidum; Strongyloides stercoralis; and Entamoeba histolytica. A donor pool was created, and screening was repeated every 4 months. Before donation, another questionnaire was used to screen for recent illnesses.
Feces were collected by the donor on the day of infusion and immediately transported to the hospital. Feces were diluted with 500 ml of sterile saline (0.9%). This solution was stirred, and the supernatant strained and poured in a sterile bottle. Within 6 hours after collection of feces by the donor, the solution was infused through a nasoduodenal tube (2 to 3 minutes per 50 ml). The tube was removed 30 minutes after the infusion, and patients were monitored for 2 hours. For patients who had been admitted at referring hospitals, the donor-feces solution was produced at the study center and immediately transported and infused by a study physician.
Eu, Procarioto
Certa vez atendi uma paciente no ambulatório do hospital e solicitei a ela, entre outros exames, um protoparasitológico de fezes. No retorno, os resultados não mostravam nada digno de nota, exceto a presença de Entamoeba coli detectada no fatídico exame. Como se sabe, esse protozoário vive de forma amistosa no organismo humano e não é causador de doenças (diferentemente de seu primo a Entamoeba histolytica). Disse a ela, então, que estava tudo bem e que poderia continuar com a medicação atual. Ela, indignada, perguntou se eu não iria tratar “aquilo”, apontando o exame com o indicador e uma cara de nojo. Eu repeti que não era necessário. Ela insistiu: “Dr. Eu não quero isso dentro de mim. Pode tratar…”
Escherichia coli
Mal sabia ela – e eu também – que há muito mais coisas nos intestinos – e no nosso organismo, de forma geral -, do que todo nosso conhecimento recente poderia supor. Não só amebas boazinhas, mas também uma infinidade de bactérias vivem em nós. Muitas bactérias. Aliás, mais bactérias que células constituintes (sim, alguém já fez essa conta!): em um indivíduo normal, existem aproximadamente 10 bactérias para cada célula humana. Isso significa que 90% das células presentes em nosso organismo pertencem a outro domínio biológico ou super-reino chamado procariotas. Número certamente suficiente para causar uma crise de identidade em minha “insegura” paciente e me fazer lembrar de Asimov no título do post.
Os estudos prosseguiram. A quantidade de bactérias abrigadas no corpo humano era tão supreendentemente gigantesca que os cientistas começaram a utilizar o nome microbioma ou microbiota, aludindo a uma possível interação ecológica entre os seres envolvidos e isso virou um projeto do Instituto Nacional de Saúde dos EUA em 2007. Ao estudar pessoas de vários lugares do mundo, descobriu-se que os respectivos microbiomas tinham diferenças significativas, tanto de pessoa para pessoa, como entre pessoas nas diversas regiões do globo. Isso lembrou os estudos “ecogenômicos” iniciados no final da década de 90. Ecogenômica, Genômica Ambiental ou Metagenômica foram nomes dados para o sequenciamento genético e identificação de microrganismos em seu habitat natural, permitindo a identificação de várias espécies que não eram vistas nas culturas clonais realizadas até então.
A presença de um “meta-organismo” geneticamente distinto dentro de nosso organismo começou a gerar perguntas sobre como seria a interação, leia-se troca de informação, entre os dois sistemas genéticos bastante diferentes e passamos a ser considerados seres metagenômicos (ou superorganismos, como preferem alguns autores) no sentido ecológico mesmo do conceito. Mas, se a “distinção galtoniana entre a genética e o meio ambiente como mecanismos geradores de nossas características fenotípicas é considerada hoje uma dicotomia simplista e (…) o meio ambiente e os genes podem interagir de múltiplas maneiras diferentes desafiando a noção de que possam agir indepententemente um do outro”, como afirma Joseph Loscalzo, editorialista do New England Journal, então, a presença desse riquíssimo material genético interagindo com o nosso deve provocar algum tipo resposta. Para descrever esse novo modo de interação, o modelo de relação hospedeiro/parasita já não parece ser suficiente porque as mesmas bactérias que nos ajudam em determinados momentos, podem nos prejudicar em outros.
Surge então, um novo mecanismo fisiopatológico. Algo com o qual não nos tínhamos defrontado antes e que, para além de quaisquer dualismos, reside na interação entre duas “forças” viventes. Antes de vencer o inimigo é preciso, agora mais do que nunca, aprender a conviver com ele.
Pflughoeft, K., & Versalovic, J. (2012). Human Microbiome in Health and Disease Annual Review of Pathology: Mechanisms of Disease, 7 (1), 99-122 DOI: 10.1146/annurev-pathol-011811-132421
Loscalzo, J. (2013). Gut Microbiota, the Genome, and Diet in Atherogenesis New England Journal of Medicine, 368 (17), 1647-1649 DOI: 10.1056/NEJMe1302154
Recomendo os textos do Meio de Cultura sobre o assunto. Superorganismos 1, 2 e 3. E o especial da Nature (em inglês, para assinantes).
Agradecimentos ao Luiz Bento do Discutindo Ecologia pela revisão do manuscrito.
O (Infra)Vermelho e o Branco
 O professor Miguel Nicolelis e sua equipe da Duke University Medical Center
O professor Miguel Nicolelis e sua equipe da Duke University Medical Center conseguiu conseguiram fazer com que ratos “visualizassem” luz em comprimentos de onda na faixa do infravermelho, o que normalmente não é possível, nem para esses animais, nem para nós. De fato, o título da matéria divulgada afirma que os roedores adquiriram a habilidade de “tocar” a luz infravermelha após a colocação de uma neuroprótese. O experimento consistia em treinar ratos a escolher cores (luz visível portanto) dentro de uma gaiola com objetivo de receber uma recompensa. “Depois de treiná-los, os pesquisadores implantaram nos cérebros dos ratos um arranjo de microeletrodos com diâmetro aproximado de 1/10 de um fio de cabelo na região cortical responsável pela sensação táctil proveniente de seus bigodes. Ligado aos microeletrodos havia um detector de infravermelho. O sistema estava programado de tal forma que quando o sensor de infravermelho disparava, era gerado um impulso elétrico no córtex sensorial do rato. O sinal aumentava em frequência e intensidade conforme o animal se aproximava da luz. No início, os ratos se confundiam e coçavam o nariz. Depois de aproximadamente 1 mês de treinamento, os ratos utilizavam o focinho para localizar a fonte de infravermelho como alguém que procura algo no horizonte com a mão em aba sobre os olhos. Levavam a cabeça para frente e para trás, procurando pela intensidade do sinal. E funcionou. Um achado importante, segundo Nicolelis, foi o fato de que tal adaptação não causou uma perda da função primária – tátil – das vibrissas do rato. Uma avenida ficcional se abre diante dessa nova possibilidade. Imaginar homens com possibilidade de “ver” ondas eletromagnéticas e outros tipos de entes invisíveis aos nossos olhos é fantástico. Além disso, a possibilidade reparar lesões neurológicas (cegueira, surdez, afasias, etc), utilizando a plasticidade do sistema nervoso é uma consequência óbvia.
A coisa toda é muito interessante. Fiquei com isso na cabeça, quando no Twitter me aparece uma palestra do Augusto de Campos tentando responder à questão: “O que é a poesia?”. Nessa palestra, Campos usa uma estorinha contada por Arnold Schönberg, o “louco” do dodecafonismo na música, quando se defrontou com a pergunta “O que é a música?”. A anedota tal como Campos contou na palestra (apesar de que o interessante e bonito portal Musa Rara dá uma outra versão) é esta:
Um cego perguntou ao seu guia: — Como é o leite?
O outro: — O leite é branco.
O cego: — E o que é esse “branco”? Me dê um exemplo de algo que seja “branco“!
O guia: — Um cisne. Ele é totalmente branco e tem um pescoço longo e curvo.
O cego: — Pescoço curvo? Como é isso?
O guia, imitando a forma do pescoço do cisne com o braço, fez com que o cego o apalpasse.
O cego: “Ah! agora eu sei como é o branco…”
Se a anedota explica (ou complica) nossa compreensão do que é a música, deixo aos seletos leitores. Tudo isso foi mesmo para dizer que se alguém perguntasse a um rato stendhaliano neuroprotetizado de Nicolelis o que é o infravermelho, talvez ele respondesse: “Infravermelho? Não sei o que é. Só sei que me dá muita vontade de coçar o nariz…”
Eric Thomson, Rafael Carra e Miguel A. L. Nicolelis. February 12, 2013 in the online journal Nature Communications.
Veja a palestra de Augusto de Campos.
O Bloqueio dos Prótons
Este é o último post da série sobre as aquaporinas, um dos mais “geniais” sistemas de transporte molecular já descoberto, que contou com os posts O Poder dos Prótons e Uma Passagem para Água.
Do ponto de vista biológico, portanto, faz sentido as aquaporinas não permitirem a entrada de espécies protonizadas. Mas, do ponto de vista bioquímico isso não é tão simples. A pergunta “Como?” é a que deve ser cientificamente respondida, mas apenas recentemente estudos com simulações elucidaram os mecanismos de bloqueio dos prótons.
São três os dispositivos capazes de efetuar o bloqueio dos prótons pelas aquaporinas[1,2]. A pergunta sobre qual seria o mais importante, ainda se constitui objeto de discussão acadêmica. A figura abaixo mostra o canal de uma subunidade da AQP1 onde quatro moléculas de água (de cor mais forte, no centro) demonstram as interações com os resíduos aminoácidos da estrutura do canal. Os mecanismos são:
(a) Estreitamento. Por restrição de tamanho, pouco acima do ponto médio do canal, o poro se estreita de 8 Å para 2,8 Å (aproximadamente o diâmetro de uma molécula de água). Isso provoca a desidratação do H3O+, o que obrigaria a molécula a desfazer-se de sua carga.
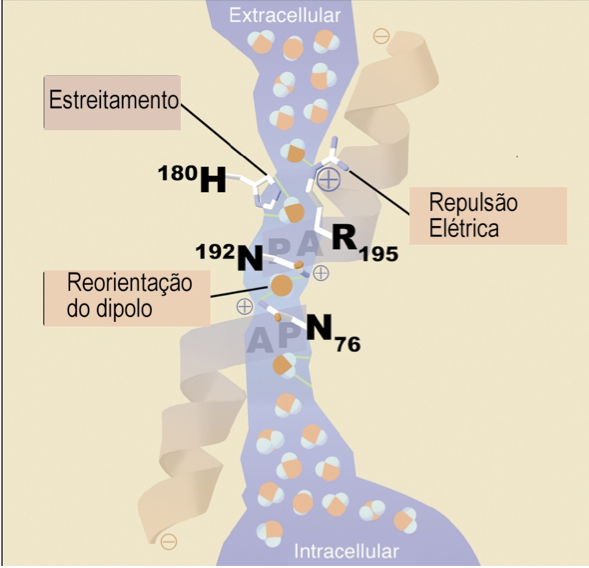 Figura 1. Mecanismos de bloqueio de prótons pelas aquaporinas. (modificado de [1])
Figura 1. Mecanismos de bloqueio de prótons pelas aquaporinas. (modificado de [1])
(b) Repulsão eletrostática. Um resíduo de arginina (R-195, na figura) na região de maior estreitamento do poro impõe uma barreira aos cátions, incluindo o H3O+. No foco da controvérsia, alguns estudos mostram que talvez esse mecanismo seja o mais importante no bloqueio dos prótons pois ele está presente virtualmente em toda família das AQP. A retirada deste resíduo reduz drasticamente a especificidade aos prótons.
(c) Reorientação do dipolo. Duas hélices parciais formadas por resíduos Asn-Pro-Ala (chamados NPA motifs) se encontram no meio do canal, formando um campo magnético bipolar que alinha a molécula de água através da formação de duas pontes de hidrogênio. Isso orienta perpendicularmente a água, fazendo com que as pontes O-H apontem para fora do canal, impedindo a condutância dos prótons pelo efeito Grotthuss. Mais elegante, esse mecanismo foi o proposto inicialmente [3,4]. A figura 2 mostra como o campo magnético bipolar exige que a molécula de água tenha uma orientação específica para entrar no canal a um baixo custo de energia.
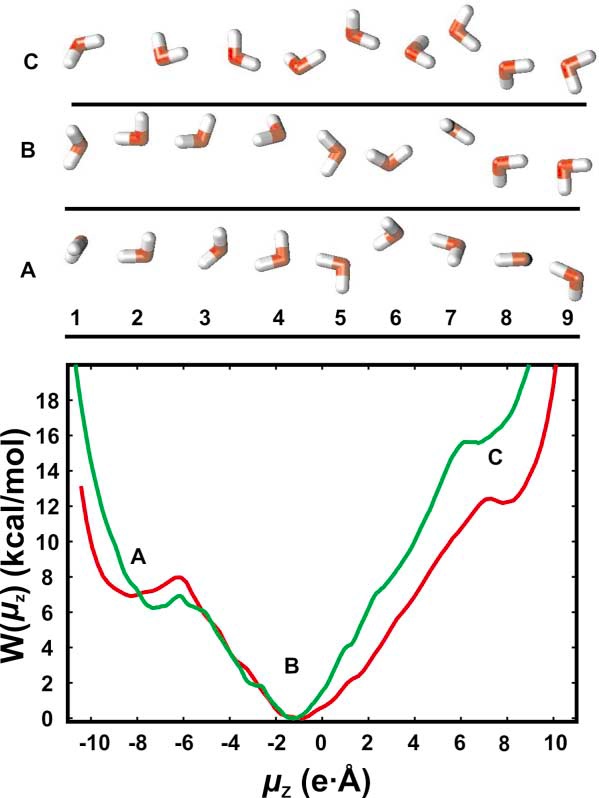
Figura 2. Modelagem da organização preferencial da cadeia de moléculas de água da aquagliceroporina (GlpF) de E. Coli. (Acima) Três conformações representativas da cadeia de água. (A) Com as pontes de H orientadas para o extracelular. (B) Conformação preferível. Note que entre a molécula 6 e a 7, o alinhamento entre H e O (branco e vermelho, respectivamente) é quebrado. (C) Com as pontes de H orientadas para o intracelular. (Abaixo) Gráfico ilustrando a energia necessária para a reorientação das moléculas de água (nas abscissas) e a orientação das pontes de H (ordenadas) por duas metodologias diferentes (linhas vermelha e verde). As letras A, B e C correspondem às conformações acima, sendo a B a de menor trabalho termodinâmico . (A partir da referência [3])
Os dispositivos de bloquear prótons presentes nas aquaporinas, seja pelo posicionamento estratégico de cargas ao longo de um canal, seja pelo “desengajamento” da corrente de moléculas de água que quebra o “teletransporte” dos prótons, são exemplos interessantes da “luta” na qual os seres vivos se envolveram para chegarmos onde estamos. Tal luta não se dá apenas no nível macroscópico da concorrência entre as espécies e a seleção natural. Ela ocorre também no nível subcelular – das moléculas -, e mostra a dificuldade de resistir e manter-se estável na agressividade do ambiente natural, o que pode bem ser entendido como “viver”. Parece, então, ter sido evolutivamente vantajoso para célula manter os sistemas de transporte de água e prótons separados de modo a poder controlar o volume e a concentração dos solutos do citoplasma, por um lado, e o metabolismo energético, por outro, de forma independente[4]. A família das AQPs é filogeneticamente antiga, estando presente nos procariotas, o que indica a urgência desse controle já nos primórdios da vida na Terra. De fato, domar precocemente as vicissitudes da água parece mesmo ter sido imprescindível para que os seres vivos prosseguissem dependendo dela.
Referências Bibliográficas
[1] Kozono D, Yasui M, King LS, et al.: Aquaporin water channels: atomic structure molecular dynamics meet clinical medicine. J Clin Invest 109:1395-1399, 2002.
[2] Chen H, Wu Y, Voth GA: Origins of proton transport behavior from selectivity domain mutations of the aquaporin-1 channel. Biophys J 90:L73-75, 2006.
[3] Chakrabarti N, Tajkhorshid E, Roux B, et al.: Molecular basis of proton blockage in aquaporins. Structure 12:65-74, 2004.
[4] Eisenberg B: Why can’t protons move through water channels? Biophys J 85:3427-3428, 2003.
Uma Passagem para a Água
Este post faz parte de uma série cujo primeiro é O Poder dos Prótons.
Na fisiologia clássica, por volta da década de 20, quando se descobriu que a membrana plasmática das células era uma dupla camada lipídica, formulou-se a hipótese de que a água pudesse penetrar no meio intracelular através da própria membrana, seguindo forças osmóticas. Entretanto, uma série de pesquisadores, por meio de medidas biofísicas (que analisam potenciais de membrana), notaram que a permeabilidade de algumas membranas à água era 10 a 20 vezes maior do que a esperada caso a passagem ocorresse apenas através delas[1]. Previram, assim, a existência de um canal para passagem da água.
No início da década de 80, Peter Agre era um hematologista envolvido no estudo do fator Rh (o “positivo” ou “negativo” dos tipos sanguíneos) trabalhando no Instituto Nacional de Saúde dos EUA. Estava interessado em induzir a formação de anticorpos em coelhos sensibilizando-os com um peptídeo parcialmente purificado do fator Rh. Os coelhos reagiam fortemente produzindo anticorpos que, no entanto, não reagiam com o cerne da molécula de Rh mas sim, com uma outra proteína que acreditava-se ser um fragmento do grande polipeptídeo. Parecia uma contaminação do experimento. Mas essa proteína de 28 kilodaltons (kDa), tinha algumas propriedades estranhas. Não se corava com os procedimentos habituais e quando foram procurá-la na membrana de hemáceas, verificou-se que era extremamente abundante. Com aproximadamente 200.000 cópias por hemácea, era uma das proteínas mais comuns na célula! Nas palavras de Agre (descendente de imigrantes nórdicos) “era como se alguém, andando pelo norte desértico da Suécia, de repente, encontrasse uma cidade de 200.000 habitantes que não constasse em nenhum mapa”. Estudos subsequentes revelaram que a proteína tinha características de um canal pois dispunha-se ao longo da espessura da membrana. Uma série de elegantes experimentos revelou que ela era um canal de água sendo batizada com o sugestivo nome de aquaporina 1 (AQP1). Peter Agre recebeu o prêmio Nobel de Química em 2003 por sua descoberta (Figura abaixo). Orgulho do pai, que havia trabalhado com Linus Pauling, acabou abandonando a hematologia. (Veja a interessantíssima “palestra do prêmio” (em inglês, 45 min) no portal do Nobel).

Figura 1. Peter Agre. Prêmio Nobel de Química de 2003
As aquaporinas constituem uma família de, até o momento, algumas centenas de proteínas de membrana. Nos mamíferos já foram identificadas doze, sete das quais estão presentes no rim. Nos últimos anos, pesquisas têm explorado a seletividade e as funções multitransportadoras das aquaporinas. Isso levou a uma divisão do grupo em aquaporinas clássicas e aquagliceroporinas. Estas últimas também transportam passivamente glicerol e outros polióis bem como alguns solutos e, ao que parece, são filogeneticamente até mais antigas que as aquaporinas clássicas[2]. A AQP1 é uma proteína de membrana altamente permeável à água. Sua condutância é de aproximadamente 3 x 10^9 molec / subunidade / seg permitindo que uma molécula de água trafegue a ~0,02 cm/s [3]. A AQP1 forma 4 complexos na membrana celular (subunidades), cada um formando um canal de água independente (figura 2). Um quinto poro é formado no centro do complexo e há indícios de que possa conduzir íons, mas o transporte passivo de água através da membrana celular é mesmo a maior função fisiológica da AQP1 [4].

Figura 2. Foto de simulação do transporte de água por uma molécula de aquaporina. As subunidades são constituídas de túbulos que estão em cores diferentes. As moléculas de água (pequenos bumerangues azuis) permeiam a subunidade à direita (em dourado). Esta imagem ganhou o prêmio de melhor foto científica da revista Science™ na edição de 24 de Setembro de 2004 e pode ser encontrada aqui, juntamente com uma animação do transporte das moléculas de água.
É surpreendente o fato de que, mesmo transportando água tão eficientemente, as aquaporinas sejam impermeáveis aos íons. A razão disso seria a proteção do meio interior celular para que a maquinaria metabólica possa funcionar adequadamente e em “silêncio biológico” – o que pode até ser uma definição bonita de Saúde, mas teleológica demais para servir a nossos propósitos. Até o mais enxerido de todos íons, o próton e seu alterego H3O+, não consegue passar pelas reentrâncias do canal de água. Como isso se dá? Como isso ocorre se os prótons teleportam-se de um lugar para outro livremente na Mattrix aquática, materializando-se e evaporando em qualquer lugar bastando para isso um “fio de água” por onde possam passar…
É o que tentarei descrever no próximo e último post da série.
Referências Bibliográficas
[1] Finkelstein A: Water Movement Through Lipid Bilayers, Pores, and Plasma Membranes: Theory and Reality (Distinguished Lecture Series of the Society of General Physiologists). New York, John Wiley & Sons Inc, 1987.
[2] Nielsen S, Frokiaer J, Marples D, et al.: Aquaporins in the kidney: from molecules to medicine. Physiol Rev 82:205-244, 2002.
[3] Preston GM, Carroll TP, Guggino WB, et al.: Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 256:385-387, 1992.
[4] Zhu F, Tajkhorshid E, Schulten K: Theory and simulation of water permeation in aquaporin-1. Biophys J 86:50-57, 2004.
O Poder dos Prótons
Há um poder, silencioso e gigantesco que, entre outras coisas, é o responsável pela água na forma como a conhecemos no nosso planeta. Se a água é, por sua vez, a matriz de todas as formas vivas daqui, então tal poder também estará presente em cada ser vivo. A explosão de vida da Terra teve que lidar com ele para que pudesse florescer e, de fato, quando nos escapa esse controle, uma fera enjaulada foge e a vida diminui ou mesmo acaba. Podemos chamar isso de “doença”. Nos próximos posts vou tentar contar a história do Poder dos Prótons e seus desdobramentos na fisiologia e fisiopatologia dos seres vivos, em especial, dos humanos.
O Hidrogênio poderia, para todos os efeitos, ser considerado um elemento-traço. Para se ter uma ideia, sua concentração é 3,5 milhões de vezes menor que a do Sódio nos fluidos orgânicos. No soro, ela é da mesma ordem de grandeza do Molibdênio (20 nM/L) e bem menor que concentrações de elementos como Zinco (15 mM/L), Cobre (20 mM/L) e Selênio (1 mM/L) [1]. Como pode o Hidrogênio com uma concentração de 0,000 000 040 M/L ou 40 nM/L influenciar tão decisivamente os processos biológicos? Quais as razões que levam o íon hidrogênio a desempenhar um papel tão importante?
A primeira delas é, sem dúvida, a água. Todos os fenômenos biológicos ocorreram e ocorrerão sempre em soluções aquosas desde que a estrutura celular que caracteriza os seres vivos neste planeta seja mantida. A água tem, como se sabe, estranhas propriedades quando comparada a outros hidretos da família do Oxigênio (a família 6A na tabela periódica – Enxofre, Selênio, Telúrio e Polônio): altos pontos de fusão e ebulição, alta constante dielétrica (conduz correntes bem), alta tensão superficial (forma gotas muito resistentes), entre outras (Tabela 1).
|
Grupo |
3A |
4A |
5A |
6A |
7A |
|
B2H6 : -92.5 |
CH4 : -164 |
NH3 : -33.4 |
H2O : +100 |
HF : -87.7 |
|
| H2S : -60.7 | |||||
| H2Se : -41.5 | |||||
| H2Te : -2 |
Tabela 1. A água tem um ponto de ebulição anormalmente elevado quando comparado por extrapolação em relação aos hidretos do grupo 6A (Oxigênio, Enxofre, Selênio e Telúrio). Os hidretos dos outros grupos são mostrados para comparação (3A Boro, 4A Carbono, 5A Nitrogênio e 7A Flúor). (Retirado daqui)
A grande maioria das estranhas propriedades da água provém da habilidade que ela tem de formar pontes de hidrogênio. Como se sabe, a ligação do Oxigênio aos dois átomos de Hidrogênio dá à molécula de água o formato de um “V” com um ângulo de 104,5o. Essa ligação é formada por um par de elétrons compartilhado (a chamada ligação covalente). O Oxigênio, no entanto, atrai o par de elétrons para bem perto de si (maior eletronegatividade) e transforma a molécula de água em um minúsculo imã com seus 2 pólos: o positivo (Hs) e o negativo (O). Quando um átomo de H, carregado positivamente, fica preso entre dois átomos de O, que têm cargas negativas, ele passa a funcionar como uma ponte entre os dois. Essa atração é 90% eletrostática e 10% covalente (há evidências de que eles podem realmente dividir orbitais, fazendo com que não consigamos saber qual ligação é a covalente), e cada molécula de água tem o potencial de fazer 4 pontes. Além disso, pode-se dizer que a ponte de hidrogênio é cooperativa isto é, uma vez formada a primeira ponte, a molécula torna-se mais apta a formar outras pontes e o fenômeno se dissemina. O contrário também é verdadeiro, ou seja, é mais difícil romper a primeira ponte, sendo que a energia necessária para romper as subsequentes vai, progressivamente, ficando menor. Isso transforma a água em um “sincício”, tipo um tecido tridimensional; daí a poderosa força de atração intermolecular que dota a água de seus elevados pontos de ebulição, fusão e tensão superficial. A formação das pontes de hidrogênio faz com que a entalpia (energia) do sistema fique mais negativa e a entropia (grau de desorganização) menos positiva ou, trocando em miúdos: a água se organiza!. Organiza-se com baixo custo, o que significa que é difícil tirá-la desse estado de organização e, na dependência da temperatura e pressão, temos a explicação de muitas das suas “estranhas” propriedades. Ou seja, se a água fosse apenas H2O seria um gás na faixa de temperatura que permite que seres vivos sobrevivam. No estado líquido, ela está na forma de (H2O)n, com n→∞, constituindo mais pontes de H que qualquer outro solvente e com quase tantas pontes quanto ligações covalentes. Esse sistema é interessante pois pode rearranjar-se rapidamente frente a estímulos como solutos ou alterações de temperatura, como uma rede tridimensional capaz de mudar o tamanho de seus “buracos” de acordo com o tamanho dos “peixes” que nela caem.
Além disso, a água demonstra uma extremamente baixa mas mensurável capacidade de formar íons. Ioniza-se devido a flutuações do campo magnético de seu dipolo causadas por vibrações complexas de sua estrutura molecular e também por posicionamentos espaciais favoráveis de suas pontes de H. Após a ionização, o H+ não permanece livre muito tempo (menos que 1% do tempo total) e é hidratado para formar o hidroxônio, oxônio ou hidrônio (H3O+). Os três átomos de H da molécula são equivalentes na ligação com o O e organizam as pontes com as moléculas de água adjacentes. O H3O+ exibe uma mobilidade frente aos campos elétricos de testes que é absurdamente maior que a prevista para um cátion monovalente (como o Lítio, por exemplo, que apresenta inclusive uma forma de hidratação semelhante). Qual seria a explicação para esse fenômeno?
Um próton pode viajar em uma solução aquosa de duas maneiras: uma, chamada hidrodinâmica, em que o H3O+ difunde-se como uma molécula comum abrindo caminho pelo meio aquoso; outra, chamada prototrópica e essa merece uma explicação. Em 1905 [2], foi sugerido que a transferência do próton poderia ocorrer por uma “rede” de pontes de hidrogênio, um processo que envolveria uma série periódica de polimerizações da água entre H9O4+ (cátion de Eigen) e o H5O2+(cátion de Zundel), conhecido como efeito Grotthuss [3], proton-wire ou proton-jumping, ou ainda water-wire. Isso só é possível, devido à efêmera duração da ponte (cada ponte tem uma duração média de 10 psec, sendo que 1 picosegundo = 10-12s) e à facilidade com que a água as forma e “re-forma”, como vimos. Dizemos então, que a alta condutividade do próton em meio aquoso ocorre devido a “mudança de identidade das moléculas de água que participam das pontes” pois os núcleos de H vão passando de uma para outra conforme esboça a figura 2, sendo que o que entra, é diferente daquele que sai.
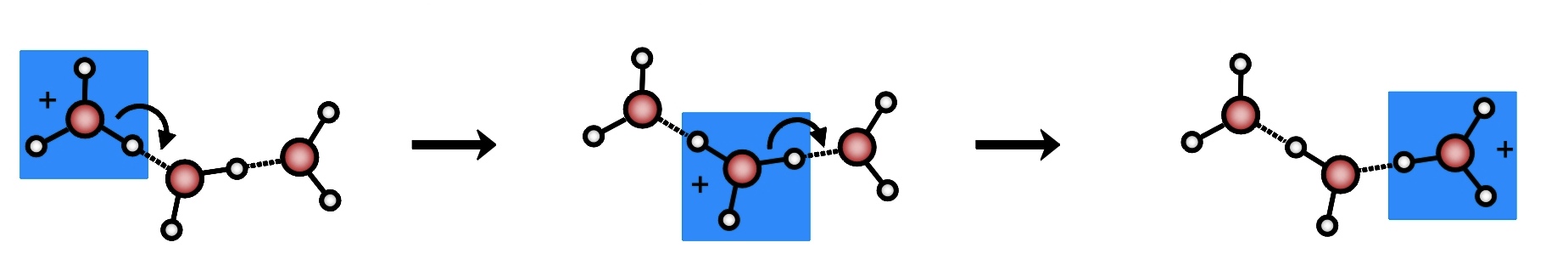
Figura 2. Esquema simplificado do efeito Grotthuss
Isso funciona parecido com o hipnotizante Pêndulo de Newton, na figura abaixo.

Pêndulo de Newton (fonte Wikipédia)
Mas…
O íon hidrogênio tem uma densidade de carga altíssima e isso ocorre porque ele é um cátion-anão (monovalente no qual não existem elétrons circulantes) sendo seu raio 105 vezes menor que qualquer outro. Por essa razão, tem um grande campo magnético ao seu redor. Assim, mesmo estando presente em concentrações baixíssimas nas soluções e devido sua alta mobilidade, o H3O+ é um “pentelho iônico” e pode afetar a conformação de proteínas, ácidos nucléicos e membranas biológicas, bastando para tal, que haja uma “brecha” eletrostática (cargas) na solução. Esse é o “poder dos prótons”, capazes de alterar todos os processos biológicos, portanto. Fenômenos semelhantes ocorrem com a hidroxila OH– entretanto, sua mobilidade nas soluções tem sido bem menos estudada. Pesquisas recentes [4] demonstram que sua mobilidade também é anormalmente alta para um ânion de seu porte e, portanto, mecanismos semelhantes ao do H3O+ podem também ocorrer. Tanto o H+ como o OH– são gerados pela destruição e destruídos pela formação de uma molécula de água. Se considerarmos que a água é, em ordem de grandeza, a mais concentrada substância dos sistemas vivos (55,3M – o que é 400 vezes mais concentrado que o sódio), e que ela provê uma fonte simplesmente inesgotável para esses íons, entenderemos porque eles se comportam de maneira tão diferente de outros íons de carga e massa semelhantes.
Com toda essa mobilidade e onipresença, é natural esperar que prótons penetrem as células com facilidade, podendo fazê-lo através da membrana ou por meio de canais, específicos ou não, para seu transporte. Tal fluxo é tão fundamental quanto o de água para a célula, pois está ligado à produção da energia que sustenta o metabolismo celular. Mas como evitar então, que o transporte de água em abundância através da membrana não cause uma catástrofe iônica na célula? Como evitar que os “pentelhos iônicos” dos H3O+ saturem as cargas das proteínas e as deformem fazendo com que percam sua função e, assim, parem o funcionamento da maquinaria celular?
É o que mostraremos no próximo post.
Referências Bibliográficas
[1] Tietz N: Clinical Guide to Laboratory Tests, 3rd ed. Philadelphia, W. B. Saunders, 1995
[2] Decoursey TE: Voltage-gated proton channels and other proton transfer pathways. Physiol Rev 83:475-579, 2003.
[3] Agmon N: The Grotthuss mechanism. Chem Phys Lett 244:456-462, 1995
[4] Tuckerman ME, Marx D, Parrinello M: The nature and transport mechanism of hydrated hydroxide ions in aqueous solution. Nature 417:925-929, 2002
Diapedese e Diálogo II
 Imagine que você está num barco no meio de um rio cuja correnteza é meio fortinha. Para conseguir atracar o barco à margem, você antes tem que se agarrar em alguma coisa fixa que possa prendê-lo apesar da força da correnteza. Só assim você conseguirá parar o barco e sair dele, pisando em terra firme.
Imagine que você está num barco no meio de um rio cuja correnteza é meio fortinha. Para conseguir atracar o barco à margem, você antes tem que se agarrar em alguma coisa fixa que possa prendê-lo apesar da força da correnteza. Só assim você conseguirá parar o barco e sair dele, pisando em terra firme.
Agora imagine um vaso sanguíneo contendo milhares de células, brancas, vermelhas e umas coisinhas pequeninhas chamadas plaquetas (ou trombócitos). O fluxo sanguíneo é rápido. Nas carótidas sem obstruções, a velocidade média do sangue medida pelo método do doppler, gira em torno dos 30 cm/s, que convertidos, fornecem o valor de 1,0 Km/h. Como o fluxo é pulsátil e muda com a posição do corpo e com o exercício, pode chegar a 300 cm/s, o que já dá é uma correntezazinha respeitável, vá! Os leucócitos são células de defesa e em muitas situações necessitam passar do interior do vaso para o tecido circunjacente. Como eles grudam na parede do vaso é que é interessante.
A imagem acima é um esboço da diapedese. A imagem abaixo é um esquema da Nature para explicá-la.
Como se pode ver pelo desenho, o leucócito é “capturado” pelo endotélio (capa de células que recobre o interior dos vasos), faz um “rolamento”, pára, gruda, rasteja e, na maior cara-de-pau, sai do interior do vaso (transmigra), seja entre as células (paracelular) ou por dentro de uma das células do endotélio (transcelular). Não se perde nem uma gotinha de sangue (nenhuma célula vermelha) nesse processo! Os retângulos acima com siglas “hieroglíficas” representam as moléculas inflamatórias que o leucócito utiliza para realizar a “ancoragem”. Veja quantas existem! Há medicações que bloqueiam ou estimulam a grande maioria delas e que podem atuar como anti-inflamatórios ou pró-inflamatórios dependendo do caso. O filme abaixo é muito didático e mostra como o leucócito para no fluxo de sangue de um vaso para quem não acreditou que isso de fato ocorresse.

Fiz uma associação no outro post entre diapedese (do leucócito) e o diálogo (com o paciente). Diapedese quer dizer “saltar através”. O radical dia em grego formou várias palavras médicas como diálise, diabetes, diafragma, diáfise, entre outras. Diálogo, bem isso já é bem mais complexo.
 * * *
* * *
Sobre Fatos
 “Me ensinaram que eu tenho um destino. Não sabiam que eu era uma fatalidade.”
“Me ensinaram que eu tenho um destino. Não sabiam que eu era uma fatalidade.”
@millorfernandes [no Twitter]
Achei esse texto apropriado às discussões recentes, a saber, o de entender algumas diferenças entre a interpretação dos fatos quando dentro de um contexto que tem o homem como objeto. É um pequeno excerto de um livro bastante interessante chamado “Trem Noturno para Lisboa” de Pascal Mercier [Record, 2009. Tradução de Kristina Michahelles]. Ainda comentarei mais sobre esse livro.
As Sombras da Alma
As histórias que os outros contam sobre nós e as histórias que nós mesmos contamos – quais delas se aproximam mais da verdade? É tão certo que sejam as próprias histórias? Somos autoridades para nós mesmos? Mas não é essa a questão que me preocupa. A verdadeira questão é: existe, nessas histórias, alguma diferença entre certo e errado? Nas histórias sobre coisas exteriores, sim. Mas quando tentamos compreender alguém em seu interior? Esta viagem algum dia chega a um fim? Será a alma um lugar de fatos? Ou seriam os supostos fatos apenas uma sombra fictícia das nossas histórias?
“Ou seriam os supostos fatos apenas uma sombra fictícia das nossas histórias?” é de uma crueza nietzscheana. O pequeno parágrafo junta teorias psicanalíticas, a formação do “eu” e a fundamentação da moral no sujeito. Qualquer consulta médica que se preze começa com o que se convencionou chamar anamnese (literalmente, não não-lembrar). Um paciente produz uma história sobre seu sofrimento, mas o médico deve estimulá-lo a produzir uma narrativa sobre si. Seu sofrimento não faz sentido se não for assim. Mas, seres humanos são bons para falar de coisas, não deles mesmos. A descrição formal das “coisas” permitiu o aparecimento da ciência. As descrições de si, são “loucuras” psicanalíticas, metade pseudociência, metade religião, com linguagem rebuscada e imprecisa, que frequentemente não chegam a lugar algum. É mais fácil falar dos outros.
O escândalo da vida é um fato. O aparecimento da consciência, uma fatalidade. Millôr sabe das coisas.
Foto: L’ombra burlona de ita145117


